Article Text

Abstract
Intestinal microbiome dysbiosis has been consistently described in patients with IBD. In the last decades, Escherichia coli, and the adherent-invasive E coli (AIEC) pathotype in particular, has been implicated in the pathogenesis of IBD. Since the discovery of AIEC, two decades ago, progress has been made in unravelling these bacteria characteristics and its interaction with the gut immune system. The mechanisms of adhesion of AIEC to intestinal epithelial cells (via FimH and cell adhesion molecule 6) and its ability to escape autophagy when inside macrophages are reviewed here. We also explore the existing data on the prevalence of AIEC in patients with Crohn’s disease and UC, and the association between the presence of AIEC and disease location, activity and postoperative recurrence. Finally, we highlight potential therapeutic strategies targeting AIEC colonisation of gut mucosa, including the use of phage therapy, bacteriocins and antiadhesive molecules. These strategies may open new avenues for the prevention and treatment of IBD in the future.
- chronic ulcerative colitis
- inflammatory bowel disease
- crohn’s disease
- e. coli
- bacterial adherence
Statistics from Altmetric.com
Introduction
IBDs, comprising Crohn’s disease (CD) and UC, are chronic idiopathic inflammatory GI disorders. The pathogenesis of CD and UC is likely multifactorial and includes genetic susceptibility factors, priming by the enteric microbiota and immune-mediated intestinal injury.1 2
Intestinal microbiome dysbiosis, characterised by an increase in the number of mucosa-associated bacteria and a reduction in the overall biodiversity, has been consistently described in patients with IBD.3–5 There is a decrease in beneficial bacteria, such as those of the Firmicutes phylum,6 7 and an increase in certain members of Enterobacteriaceae, thought to play a pathogenic role in disease development.8 9 Although no single causative microorganism has been identified, many studies have reported the potential involvement of microbial pathogens (Mycobacterium avium paratuberculosis, 10 11 Clostridium difficile,12 Helicobacter species,13 Campylobacter species)14 15 and functionally altered commensal bacteria (invasive Escherichia coli, 16 17 enterotoxigenic Bacteroides fragilis)18 in disease pathogenesis. The frequent recovery of E coli adhering to the inflamed ileal and colonic mucosa of patients with CD19–24 and UC25 26 has stimulated great interest over these bacterial species in the past 15 years. The high prevalence of adherent E coli bacteria isolated from the ileal mucosa of patients with CD led to the characterisation of several strains, which failed to detect any virulence factor-encoding genes that are traditionally present in typical pathogenic species.20 One property of these strains is the ability to adhere to and invade intestinal epithelial cells (IEC), as well as to replicate within macrophages; hence they were categorised as a new specific pathogenic group of E coli called adherent-invasive E coli (AIEC).27 28 Several molecular and culture-based investigations support a putative role of AIEC in CD; however, controversy exists regarding its role in the pathogenesis of UC.16 20 26 29 This review aims to provide an update on the potential role of AIEC in the pathogenesis of IBD based on a systematic search of the literature. We also explored strategies targeting AIEC colonisation of gut mucosa as a potential therapeutic option for patients with IBD.
Methodology for literature search and selection criteria
A comprehensive literature search was performed to identify all relevant citations that have reported AIEC in IBD. We conducted a systematic search on PubMed and Scopus database using the following keywords ‘Escherichia coli’[Mesh] OR Escherichia coli AND adherent AND ‘Inflammatory Bowel Diseases’[Mesh] OR Inflammatory Bowel Diseases OR IBD OR Inflammatory Bowel Disease OR Ulcerative Colitis OR Crohn Disease OR Crohns Disease OR Crohn’s Disease OR Crohn’s Enteritis OR Regional Enteritis OR Granulomatous Enteritis OR Ileocolitis OR Granulomatous Colitis OR Terminal Ileitis OR Regional Ileitides OR Regional Ileitis. We restricted this search to English-language papers published up to April 2017. A total of 212 original papers were retrieved. This search was complemented by a manual review of the reference list of included studies as well as relevant review articles of the topic. Studies were included if they fulfilled the following criteria: (1) original investigation characterising the AIEC pathotype, the different strains and virulence properties; (2) original studies reporting AIEC interaction with epithelial barrier cells and/or macrophages; (3) original papers addressing AIEC and E coli prevalence in IBD; (4) original articles that have reported AIEC in IBD postoperative recurrence or disease activity or (5) original papers or review articles that have discussed the possible role of AIEC in IBD pathogenesis and any potential therapeutic interventions. After selection, a total of 214 original papers were included.
Definition of AIEC
AIEC strains are considered to be pathobionts because they promote inflammatory diseases due to the adaptive evolution of their genome in a specific and susceptible host.30–33 In contrast to opportunistic pathogens, pathobiont influence on the host occurs indirectly via stimulation of the immune system.34 The AIEC phenotype is associated with (1) the ability to adhere to IEC,20 (2) the ability to invade IEC through the involvement of host cell actin polymerisation and microtubule recruitment,27 (3) the ability to survive and replicate within macrophages without inducing cell death35 and (4) the absence of any specific known invasive determinants.17 To date, assessing the interaction between bacterial and host cells is the only way to efficiently identify AIEC strains. Although molecular approaches and genomic sequencing have identified genes associated with AIEC invasion of IEC and replication in macrophages, a specific molecular marker linked to the AIEC pathovar is still missing.
AIEC strains harbour genetic similarity, in terms of phylogenetic origin and virulence genotype, with extraintestinal pathogenic E coli (ExPEC). Only 6.3% of ExPEC strains are AIEC based on their phenotypic features.36 AIEC has been isolated from different populations including adults and children,33 37 healthy subjects, patients with IBD or colorectal cancer,29 companion animals, such as boxers with granulomatous colitis,38 and animals with bovine mastitis,39 suggesting the AIEC pathotype to be disease rather than host specific.
Different AIEC strains
Pulsed-field gel electrophoresis, multilocus sequence typing, PCR-based typing analysis, core-genome-based phylogenetic analysis and serotyping have shown that AIEC strains are clonally diverse and belong to distinct phylotypes and serotypes. Although they are often assigned to the B2 phylogroup, which contains the most virulence factors,40 strains belonging to the A, B1 and D phylogroups have also been isolated.16 22 33 41–46 This phylogenetic distribution suggests that the AIEC phenotype has likely evolved several times.
One of the first AIEC strains isolated in 1998 from the ileum of patients with CD,20 named LF82 (for Lille-France), serves as a reference for most studies involving IBD-associated E coli strains. The complete genome of this prototype strain has been reported,31 as well as that of three other highly invasive human AIEC isolates NRG857c,30 UM14647 and HM605,48 which all belonged to the B2 phylogroup. LF82 and NRG857c strains appear genetically related as both belong to the O83:H1 serotype, B2 phylogroup and ST135 sequence type (ST). However, other AIEC isolates, such as 541-1, 541-15 and 576-1 sequenced strains,16 45 represent diverse serotypes with a large range of ST and phylogenetic lineages.30 43 45 49 Comparative genomics carried out in different AIEC strains and other pathogenic and non-pathogenic E coli have shown high genotypic variability, making the identification of AIEC-specific genetic factors challenging. Nonetheless, AIEC resembles ExPEC in diseases such as urinary tract infections (uropathogenic E coli), neonatal meningitis (neonatal meningitis E coli) and avian colibacillosis (avian pathogenic E coli) in terms of virulence factors.30 36 43 50 51 AIEC genomes are enriched in genes belonging to the type 6 secretion system, which could allow for the transfer of effector proteins to kill adjacent bacteria or act on host cells to facilitate intracellular survival30 31 52; however, most of them are part of B2 phylogroup.43 Likewise, genes encoding propanediol utilisation proteins, specifically the pdu operon, are significantly correlated with the AIEC pathotype and rarely present in other E coli strains.45 49 52 Recently, we reported that a subpopulation of AIEC bacteria is genetically equipped to colonise Peyer’s patches (PPs), which may ensure their rapid dissemination in the intestinal mucosa and to the mesenteric lymph nodes (MLNs). The evaluation of patient stools for E coli strains carrying gipA and lpfA genes could constitute an efficient test to identify a subset of patients with CD that may benefit from therapeutic elimination of AIEC.53 54 However, while controversial, clustered regularly interspaced short palindromic repeat (CRISPR) analysis in comparative transcriptomics should be completed to determine whether particular CRISPRs are associated with the AIEC phenotype.49 52
Pathogenic features of AIEC
It is unclear whether AIEC bacteria trigger intestinal inflammation leading to IBD or whether they serve as an aggravating factor by colonising the mucosa of patients with pre-existing inflammatory disease. Several factors involved in the persistence of AIEC in the gut mucosa and therefore promoting inflammation have been identified. The absence of conventional pathogenicity genes suggests an important role for host defects, such as abnormal mucosal immunity (particularly in CD) or intestinal barrier dysfunction (particularly in UC), contributing to AIEC’ s ability to invade mucosal cells.55 56 AIEC virulence genes can promote motility, capsule and lipopolysaccharide (LPS) expression, serum resistance, iron uptake, adhesion to and invasion of epithelial cell lines and biofilm formation, with AIEC strains containing bacterial elements and regulatory pathways that modulate these key pathogenic determinants. For example, lipoproteins,57 58 histone-like proteins,59 ribonucleotide reductase NrdR,60 the flagellar transcription regulator FlhD2C2, sigma factor FliA and the second messenger cyclic dimeric GMP (c-di-GMP)61 are involved in interactions with IEC, while the sigma(E) pathway is implicated in biofilm formation.62–64 Additionally, the ability to form biofilms on IEC65 as well as long polar fimbriae (LPF) are pathogenic features and properties of AIEC (table 1).54
Potential virulence factors described in adherent-invasive Escherichia coli pathobiont
Other AIEC virulence properties are linked to their survival and replication within macrophages. For example, the LF82 strain does not escape into the cytoplasm but replicates in mature phagolysosomes, despite the acidic pH, oxidative stress, proteolytic enzymes and antimicrobial compounds.66 AIEC survival and replication in macrophages is promoted by the stress protein HtrA,67 the thiol-disulfide oxidoreductase DsbA,68 the RNA-binding protein Hfq69 and the FAD-dependent oxydoreductase IbeA70 that play a role in surviving reactive oxygen species (ROS) (table 1). In response to AIEC infection, tumour necrosis factor α (TNF-α)35 71 72 and other proinflammatory cytokines73 are released, further enhancing dysbiosis and proliferation of AIEC without inducing host cell death. AIEC are able to exploit host mechanisms of apoptosis by increasing S-nitrosylation and proteasomal degradation of caspase-3 in infected macrophages,74favouring their own intracellular replication through modulation of the ubiquitin proteasome system in infected-IEC,75 76 and through the release of exosomes from infected cells.77 In addition, another AIEC property is induction of autophagic neutrophil cell death.78 In the presence of antibiotics, AIEC activate neutrophil ROS production and contribute to intestinal inflammation.79 Finally, AIEC also have the ability to evade the host immune response by suppressing interferon γ (IFN-γ)-mediated signal transducer and activator of transcription-1 phosphorylation in IEC, preventing an appropriate antimicrobial response.80
AIEC and the gut immune system
AIEC strongly adheres to and colonises IEC before translocating and entering the submucosal compartment. During intestinal inflammation, the mucosal barrier is disrupted and specific receptors in the epithelium are abnormally expressed, leading to enhanced intestinal permeability and AIEC pathogenesis.81–85 Figure 1 and online supplementary file 1 summarise the different interactions of AIEC with the intestinal mucosa in IBD.
Supplementary file 1

Interaction of adherent-invasive Escherichia coli (AIEC) with intestinal mucosa in the context of IBD. (1) AIEC crossing the mucous layer and resisting antimicrobial peptides; (2) AIEC adhering to intestinal epithelial cells (IECs) and leading to colonisation of the gut mucosa; (3) AIEC entering lamina propria and Peyer’s patches via M cells and interacting with immune cells.
AIEC, adherent-invasive escherichia coli; DC, dendritic cell; GC, gobelet cell; IECs, intraepithelial cells; IEL, intraepithelial lymphocyte; IESC, intraepithelial stem cell; LT, lymphocyte T; M cell, microfold cell; NT, neutrophil; PC, paneth cell.
Interaction with epithelial cells
Intestinal E coli pathovars most likely penetrate the mucous layer by promoting mucin degradation with proteases.86 87 A recently described protease secreted by AIEC (VAT-AIEC) has been reported to contribute to gut colonisation in a murine model by enhancing the spread of bacteria through the mucous layer and adhesion to IEC (figure 1).88 The secretion of host defence antimicrobial peptides can also be altered by some isolates of AIEC by the presence of two genes in a plasmid-encoded genomic island (called PI-6), arlA, which encodes a Mig-14 family protein implicated in defensin resistance, and arlC, an OmpT family outer membrane protease.89
Adhesion to intestinal epithelium is mediated by type 1 pili (FimH) expressed on the surface of AIEC isolates via interaction with cell adhesion molecule 6 (CEACAM6) receptors on ileal enterocytes (figure 1).90–93 Polymorphisms in the fimH type 1 pilus subunit (adhesin tip protein) sequence have been reported in AIEC strains, conferring higher adhesion ability.46 94 CEACAM6 host receptors are overexpressed at the apical surface of IEC in patients with CD, thus enhancing the colonisation of ileal mucosa by AIEC.90 95 Additionally, LF82 and 083:H1 strains are able to increase CEACAM6 and LAMP1 expression on the surface of IECs, and increase HLA-DR and ICAM1 expression in the lamina propria.96 Some factors expressed by IEC, such as the protective protease meprin, impair AIEC binding to mannosylated host receptors by degrading type 1 fimbriae in a dose-dependent manner, thus protecting ileal mucosa.97 Decreased levels of meprin noted in patients with CD are believed to increase production of the proinflammatory cytokine interleukin-8 (IL-8) and thus promote AIEC colonisation.97 This colonisation promotes the expression of the pore-forming tight junction protein claudin-2 in IEC98 and a reorganisation of tight junctions from the apical side by displacing zonula occludens-1 and E-cadherin, leading to a decrease and loss of barrier function.81 99 100 When interacting with the epithelial barrier, AIEC, unlike non-motile strains, forms biofilms. Those with the H1 flagellar antigen are the strongest biofilm producers.65 Flagella on the AIEC surface are able to activate and upregulate toll-like receptor 5 (TLR5), Nod-like receptor family member NLRC4 (previously named IPAF), and membrane and cytosolic flagellin receptors, raising the inflammatory mucosal immune response in dextran sodium sulfate (DSS)-treated mice.101 The binding of flagella to TLR5 on IEC induces the production of hypoxia-inducible factor-1a and activates the nuclear factor-kappa B (NF-κB) pathway and IL-8 secretion. This leads to an increased inflammatory response and an inhibition of the autophagic process of the host cells.102–105
AIEC’s outer membrane vesicles (OMVs), via the transmembrane protein OmpA, have the ability to interact with the endoplasmic reticulum stress response factor Gp96 on IECs57 106 107 and, in association with flagellin that target TLR5, elicit an epithelial IL-8 inflammatory response105 and promote internalisation into the intestinal mucosa (figure 1). Darfeuille-Michaud’s team constructed several isogenic mutants that highlight the decreased ability of LF82 to adhere to and invade Intestine-407 cells via a downregulation of type 1 pili and flagella in nlpI-negative mutants,58 through OMV release in yfgL-negative mutants,57 through the sigma(E) regulatory pathway in ompC-negative isogenic mutants62 or by interacting with the host molecule Gp96 in ompA-negative mutants.107
Finally, FimH can bind M cells, which are distributed among enterocytes, through the recognition of apical glycoprotein 2.108 Highly glycosylated CEACAM receptors expressed on the apical membrane of M cells have been proposed as potential receptors for microbial adhesion.109 M cells account for about 5% of cells in the follicle-associated epithelium overlying PPs, are the site of aphthoid ulcers in patients with CD110 and are structurally and functionally specialised for transepithelial transport, delivering foreign antigens and microorganisms to organised lymphoid tissues within the mucosa.111 Several invasive bacteria, including Yersinia, Salmonella, Listeria and Shigella spp,112 113 use M cells to cross the gut barrier. Similarly, AIEC are able to enter and translocate through M cells and isolated human ileal PPs via a mechanism involving LPF expression (figure 1).114–117
Interaction with immune cells
Macrophages are a key component of the intestines innate immune defence by limiting systemic microbial dissemination through phagocytosis, sensing and responding to microbial stimuli and informing consequent host immune responses.118 Through pattern recognition receptors, macrophages recognise the conserved microbial molecular patterns synthesised by resident and pathogenic intestinal bacteria, including extracellular structures, such as fimbriae, flagella, LPS and peptidoglycan. Environmental factors, such as iron availability, influence the microbial production of some of these extracellular structures, including curli fibrils in Salmonella enterica serovar typhimurium 119 and type I fimbriae in E coli,120 resulting in environmental modulation of microbial interactions with macrophages.
AIEC have been immunodetected in the macrophages in germinal centres of lymph follicles of patients with CD and are the dominant species cultured from MLN of patients with CD.121 122Monocyte-derived macrophages (MDMs) from patients with CD are unable to restrict intracellular AIEC compared with healthy controls (HCs) in vitro, leading to a disordered inflammatory response influenced by disease activity.123 124 E coli possesses four known acid resistance systems. The first system requires sigma factor RpoS and the cyclic AMP receptor protein CRP, with RpoS functioning as a major environmental stress response regulator in both E coli and Salmonellae. 125 Deletion of RpoS from AIEC (strain O83:H1) isolated from subjects with CD has been observed to increase the sensitivity of this clinical isolate to oxidative stress.126 The second system requires extracellular glutamate. The components of the glutamate-dependent acid response are two isoforms of glutamate decarboxylase encoded by gadA and gadB, and a glutamate-γ-aminobutyric acid antiporter encoded by gadC.127 128 Murine AIEC have been observed to respond to chronic intestinal inflammation by upregulating expression of gadA and gadB. 129 The third acid resistance system requires an arginine-dependent utilisation of the arginine decarboxylase (AdiA and AdiC) antiporter,125 while the fourth is lysine dependent, involving lysine decarboxylase.128 In addition, E coli also harbours specific mechanisms that enable them to resist high levels of ROS that form the oxidative and superoxidative response to phagocytized pathogens. These defensive resources have recently been found to be grouped into two regulated sets of genes, soxRS and oxyR regulons.130 131
Key genes support AIEC survival and replication within macrophages. One particular gene that contributes to AIEC’s ability to survive and replicate inside macrophages is gipA. GipA expression is induced by certain intracellular, GI (bile salts) and phagolysosomal (ROS and acid pH) conditions. Deletion of gipA dampens the proinflammatory response and AIEC replication inside macrophages.53 AIEC killing by macrophages could be inhibited by microbial mannan in a TLR4 and MyD88-dependent manner.132
The LF82 strain is able to invade and replicate within human neutrophils, but in contrast to its behaviour inside macrophages and IEC, induces the autophagic death of infected neutrophils. These cells later undergo an alternative cell death process called NETosis.78 Additionally, LF82 was able to replicate within monocytes isolated from patients with CD for the first 20 hours after infection before being cleared.133 Interestingly, these cells had a reduced capacity to support the expansion of allogeneic Th17 cells from CD4+ memory T cells.134 The authors proposed that mucosal Th17 activation in patients with CD is a secondary event in response to poor bacterial clearance due to defects in innate immunity.134 Viladomiu et al revealed that AIEC isolated from patients with CD with spondyloarthritis induce Th17 mucosal immunity in germ-free mice, as well as specific pathogen-free mice, compared with non-AIEC strains. This demonstrates that AIEC can induce Th17 polarisation, even in the presence of other commensal microbiota, which requires the virulence-associated metabolic enzyme propanediol dehydratase.135 Recently, it has been shown that infection with the LF82 strain could induce the release of exosomes, which activate NF-κB, mitogen-activated protein kinases p38 and c-Jun N-terminal kinase, as well as increase the secretion of proinflammatory cytokines in vitro and in vivo. This suggests that exosomes are mediators of the host–AIEC interaction and are able to activate the innate immune response and subvert the control of AIEC replication.77 Extracellular microbial structures that impact interactions with macrophages are also produced within multicellular microbial communities, including biofilms and bacterial aggregates. Cells recovered from bacterial aggregates are more susceptible to phagocytosis since disrupted cellulose production prevents aggregation, reducing uptake of the AIEC strain NC101 by macrophages. Iron promotes the cellulose-dependent aggregation of NC101 with abrogation of bacterial cellulose production delaying the onset of colitis in inflammation-prone IL-10 -/- mice mono-associated with NC101.136
AIEC and cytokine production
LF82 and 083:H1 strains induce increased expression of TNF-α, IFN-γ and IL-8 transcripts on colon biopsies of patients with CD and affect cell cycle distribution on Caco2 cell lines.96 AIEC-infected MDM from patients with quiescent CD release significantly higher amounts of IL-6 and TNF-α than those with active disease or from HC.123 124 LF82 can activate NF-κB signalling in IEC through IκB-α phosphorylation, NF-κB p65 nuclear translocation and TNF-α secretion. Unlike Salmonella, LF82 bacteria were unable to activate caspase-1 and induce IL-18 production.76 In addition, AIEC bacteria modulate the turnover of the ubiquitin proteasome system in infected IECs by downregulating the NF-κB regulator CYLD, leading to IκB-α degradation and NF-κB activation. Flagella are able to elicit the secretion of the proinflammatory cytokine IL-8 and chemokine CCL20 in polarised IECs, which in turn leads to the recruitment of macrophages and dendritic cells to the site of infection.104 105 The secretion of IFN-γ and TNF-α by macrophages and lymphocytes leads to CEACAM6 expression, which enhances AIEC colonisation.
AIEC in IBD
AIEC in CD
Genetic and environmental factors associated with the presence of AIEC
Multiple defects in the innate immune response associated with CD could promote AIEC-induced inflammation.56 Autophagy-related risk polymorphisms lead to impaired sensing and handling of intracellular bacteria by the innate immune system, resulting in inflammation.137 138 Autophagy is induced in host cells on AIEC infection and is required to restrain its intracellular replication.71 139 140 CD-associated nucleotide-binding oligomerization domain-containing-2 (NOD2) mutations have been linked to autophagy via their interaction with ATG16L1 in response to bacterial infections.141–143 NOD2/CARD15 mutations are considered to be loss-of-function mutations with evidence of reduced production of antibacterial defensins by Paneth cells and a reduced IL-8 response to muramyl dipeptide by macrophages.144 Exposure to AIEC strains results in an increase of LC3 and ATG16L1 in NOD2 overexpressing Caco2 cell lines compared with wild-type Caco2 cells. This is accompanied by a reduced AIEC survival rate and decreased cytokine expression.145 The intermediate filament vimentin, expressed on the surface of mesenchymal cells, has been recently proposed to act as a receptor for AIEC.146 On the intracellular side, vimentin leucine-rich repeats interact with NOD2, leading to the recruitment of these proteins to the plasma membrane. This is necessary for the proper function of NOD2 in antigen detection, NF-κB activation and autophagy induction. Patients with CD have specific NOD2 variants (L1007fs and R702W) that are unable to interact with vimentin and, in turn, localise in the cytosol. This leads to a defective inflammatory response, autophagy induction and mishandling of CD-associated AIEC.146 147 Altogether, NOD2 and vimentin appear to play an important role in AIEC recognition. Polymorphisms in these two proteins may have an impact on the ability of AIEC to colonise the host.
Knock-down of ATG16L1 and IRGM results in defective recognition and clearance of Crohn’s mucosa-associated E coli within IEC and macrophages.139 However, deficiency in either gene does not interfere with the replication and survival ability of other non-pathogenic, environmental, commensal or gastroenteritis-inducing E coli, suggesting a specific role for autophagy in restraining AIEC. Similarly, expression of the Crohn’s variant ATG16L1*300A in intestinal Caco2 epithelial cells impairs their ability to capture internalised Salmonella spp within autophagosomes148 and is also associated with abnormalities in Paneth cell granule exocytosis.149 Moreover, increased levels of microRNAs miR106B and miR93 in the colon tissue of patients with active CD decrease the expression of ATG16L1,150 while overexpression of microRNA miR-196 downregulates a variant in IRGM. 151 AIEC upregulate levels of miR30C and miR130A in T84 cells and mouse enterocytes through the NF-κB pathway, leading to reduced levels of ATG5 and ATG16L1. This inhibits autophagy and enhances the inflammatory response.
As reviewed above, the adherence of AIEC to ileal IECs is also dependent on the expression of CEACAM6. In patients with CD, the overexpression of CEACAM6 and Gp96 receptors on the apical membrane of IEC enables AIEC adhesion and invasion.90 107
Studies in animal models have highlighted the importance of preceding dysbiosis and/or gut inflammation for the successful infection with AIEC. In mice, AIEC colonisation only occurs after treatment with DSS to induce colitis,101 treatment with antibiotics95 152 153 or following a high-fat and high-sugar diet.154 A plausible model for the development of CD is that alteration of the gut microbiota through an acute event, such as an infectious gastroenteritis or the use of broad-spectrum antibiotics, creates a niche for AIEC proliferation. The potentiation of acute infectious gastroenteritis in mice previously colonised with AIEC can lead to the expansion of tissue-associated AIEC and a chronic intestinal inflammatory response with subsequent bowel damage.155 These findings suggest that individuals colonised by AIEC at the time of acute gastroenteritis may be at a greater risk for CD development.155
Prevalence of AIEC worldwide in CD
Table 2 illustrates the prevalence of AIEC in subjects with CD, UC and controls from different geographical origins.
Prevalence of adherent-invasive Escherichia coli (AIEC) in patients with IBD and controls from different geographical origins
The prevalence of AIEC varies widely between studies, mostly because of small sample sizes and the use of different molecular methods of identification. There are several limitations to epidemiological studies addressing the prevalence of AIEC in IBD. First, the use of specific molecular methods to detect and quantify AIEC is highly time-consuming. The only way to efficiently identify AIEC strains is by performing assays on the interaction between bacteria and intestinal cells. Second, studies have employed different bacterial isolation methods and used different sample types.156 157 In healthy subjects, there is a significant difference between the flora found in different GI segments, but also an important variation between mucosa-associated and faecal microbiota.158 Only intestinal mucosa biopsies will contain bacteria able to adhere to IEC. Thus, studies investigating the mucosa microbiota after the removal of the mucous layer will more accurately reflect the bacteria involved in the pathogenesis of IBD than those using faecal material.29 However, in clinical practice, faecal samples are easier and less invasive to obtain. Dickinson et al 157 collected faecal samples from patients with UC to assess the incidence of AIEC and considered the methodology valid based on a previous study on colonoscopy, which revealed that the dominant serotypes in faecal samples also dominate the entire colonic flora.159 However, validation that intestinal colonisation leads to faecal shed of AIEC strains with identical characteristics is needed. Third, not all studies have used the same definition to assess adherence and invasiveness of AIEC. For instance, some authors consider an E coli strain to be adherent if the adhesion index is ≥1 bacteria per cell,17 56 while others define adherence as when the bacteria adhere to >40% of the cells.33 Lastly, studied tissue samples were heterogeneous with regards to the degree of underlying disease activity. Some studies used endoscopically active patients,56 while others included patients in endoscopic remission.29 Overall, mucosa-associated bacteria have been shown to be more abundant in inflamed than in non-inflamed IBD mucosa.56
The presence of AIEC strains in healthy individuals ranges from 0% to 16% in colonic samples and from 6% to 19% in ileal samples,17 23 41 160 meaning that these bacteria induce colitogenic effects only in combination with other CD-associated factors such as host genetic susceptibility and environmental stimuli. In healthy individuals, bacterial translocation and invasion of the mucosa is infrequently observed.161 In addition, intracellular E coli is rarely cultivated from the colonic mucosa of healthy individuals.16
Overall, the prevalence of AIEC in the mucosa of adult patients with CD ranges from 21% to 63%16 17 29 41 81 160; most investigations were undertaken in Europe and North America (table 2). However, to be considered causal in the pathogenesis of CD, its presence in patients from countries with a high or increasing incidence will need to be demonstrated.162 163
Association of AIEC with age at onset
There are few data about AIEC and age of CD onset. Studies in the paediatric IBD population reported a lower prevalence of AIEC strains compared with the adult IBD population. Negroni et al 37 found two AIEC strains from a total of 85 lactose-fermenting isolates retrieved from 34 IBD children (24 CD, 10 UC) and 23 controls. More recently, Conte et al 33characterised 616 E coli isolates from ileal mucosal biopsies from four patients with paediatric CD and four paediatric control patients. The relative abundance of AIEC strains was significantly higher in patients with CD (10% vs 1%; P<0.001). Altogether studies of AIEC in paediatric and newly diagnosed patients with IBD22 have provided further evidence for its importance in the early stages of IBD; no data are available regarding AIEC and elderly onset CD.
Association of AIEC with CD location, disease activity and postoperative recurrence
AIEC strains are more likely to be found in ileal CD than colonic CD. Darfeuille-Michaud et al 17 found that AIEC strains more frequently in the ileum (36.4% early lesions, 21.7% chronic lesions and 6.2% controls) than in the colon (3.7% of patients with CD and 1.9% of controls) of patients with CD. Martinez-Medina et al 41 164 reported a higher prevalence of AIEC in patients with ileal CD (66.7% of ileal and 58.3% of colonic samples) than those with colonic disease (50% of ileal and 25% of colonic samples). Studies have reported differences in the genetic susceptibility and immune response of the enteric microflora based on disease location,165 implying that the composition and distribution of the mucosal microbiota may also vary according to disease phenotype. Most studies lacked information on the location of CD, which is needed to determine the role of AIEC in relation to CD phenotype.
There are some data supporting the role of E coli infection on disease activity and severity, though specific data on AIEC are still lacking. Sepehri et al 22analysed right-colon biopsies at diagnosis from 34 adult patients with IBD (23 CD, 11 UC). They found that E coli isolates belonging to B2 and D phylotypes were associated with inflammation. A study by Mylonaki et al 161 using fluorescent in situ hybridisation demonstrated an increased number of E coli in the epithelium and lamina propria of patients with active CD compared with inactive disease. Likewise, other investigations found that E coli abundance was greater in patients with active CD compared with patients in remission.81 166 Elliott et al 56 found an association between the abundance of mucosa-associated E coli and the severity of endoscopic inflammation; however, this association was not found with intracellular E coli. Similarly, Baumgart et al 16analysed the mucosa-associated flora of the ileum in patients with ileal CD, colonic CD and HC. Patients with ileal CD harboured a higher number of E coli and its abundance was significantly associated with ileitis severity. More recently, Lopez-Siles et al 166 reported an association between higher numbers of E coli and a reduced interval to disease flare. These findings are in agreement with previous data reporting a correlation between increased levels of E coli outer membrane protein C (OmpC) and complicated CD with a greater need for surgery.167
Several studies have associated the presence of AIEC with ileal recurrence after surgery for CD. In their original paper, Darfeuille-Michaud et al 20 recovered E coli strains from 65% of chronically inflamed lesions (resected ileum specimens) and from 100% of the biopsies of early postoperative recurrent lesions. A significantly higher number of specimens with E coli colonisation were retrieved from early recurrent ileal lesions than from the healthy ileal mucosa of patients with CD or HC.20 These findings suggest that E coli may have a role in the initiation of the inflammatory process after surgery in CD. Nonetheless, the same authors found AIEC strains in 22% of patients with minimal or no endoscopic recurrence, suggesting that other factors are necessary to initiate CD recurrence.17 Recent data from the POP-REMIND cohort of 170 patients with CD who underwent surgery showed that the presence of AIEC on the surgical specimen was associated with severe endoscopic postoperative recurrence at 6 months (Rutgeerts score of i3 and i4).168 Targeting AIEC colonisation may be a potential adjuvant strategy to prevent severe postoperative recurrence in the future.
AIEC in UC
Predisposing factors
Controversy exists regarding the role of AIEC in the pathogenesis of UC. As discussed for CD, a defect in autophagy could favour AIEC persistence in the gut. Autophagy is blocked at the autolysosomal step following AIEC infection.78 Often during this process, pathogen-containing vacuole becomes damaged and targeted for autophagic clearance.139 Decreased levels of chromosome-associated protein D3 (CAP-D3) in patients with active UC were highlighted to impair autophagy and intracellular bacterial clearance. This is possibly a way for AIEC to regulate the recruitment of amino acid transporters implicated in CAP-D3 expression.169
Prevalence of AIEC worldwide in UC
The prevalence of AIEC in UC is less clear than in CD. Although several studies have confirmed a higher prevalence of adherent E coli strains (in some studies with a higher frequency than in CD),44 170 few have addressed the presence of AIEC bacteria in the colon of patients with UC.17 23 Most of these studies have reported a lower prevalence compared with that of CD ranging from 0% in colonic samples from adults with UC23 56 to 10% in the colons of children with UC37 81 (table 2). Several studies showed an increased abundance of E coli in active UC lesions compared with inactive mucosa or in HC.161 171 Petersen et al 172 reported a correlation between the presence of E coli from phylogenetic group B2 with at least one adhesion-related gene and increased disease activity in patients with UC (86% in active vs 13% in inactive disease). Homer et al 141 found E coli strains with adhesive or invasive properties in 35% and 27% of faecal samples from patients with active and quiescent UC, respectively (compared with 5% in controls; P<0.05). However, simultaneous adherent and invasive proprieties were not explored. Two other studies reported no increase in AIEC in patients with UC compared with HC.17 29
AIEC and IBD: primary or secondary event?
Currently it is unclear whether AIEC cause intestinal inflammation and thus could lead to disease; or whether AIEC overgrowth in the intestinal mucosa could be a consequence of pre-existing inflammation and lead to a worsening of disease.173
As outlined above, studies in animal models have highlighted the importance of preceding dysbiosis and/or gut inflammation (such as DSS colitis or antibiotic therapy) for successful infection with AIEC.101 153 These data suggest that AIEC proliferation in the gut can be seen as a consequence of inflammation.
On the other hand, several papers have confirmed the pathogenic properties of AIEC and that AIEC infection itself could induce long-term changes in the gut microbiota. Chassaing et al 174 conducted an investigation on flagellin receptor TLR5 knockout mice, which are animal models susceptible for the development of spontaneous colitis. The authors found that transient colonisation of T5KO mice by AIEC induced intestinal dysbiosis, with a greater proinflammatory potential due to increased levels of LPS and flagellin. These changes to the microbiota could cause chronic inflammation and disease. In a recent study by Small et al, 152 persistent infection with AIEC strains in streptomycin-treated mice was associated with CD-like symptoms. Additionally, the presence of intracellular E coli in the uninflamed mucosa in CD indicates that invasion by E coli may not be dependent on the presence of an inflamed and disrupted mucosal barrier.42 These results suggest that mucosal invasion by E coli may be an early pathogenic event.56 Overall, these data suggest that AIEC proliferation in the gut can also be seen as a cause of inflammation.
Current and future strategies targeting AIEC in IBD
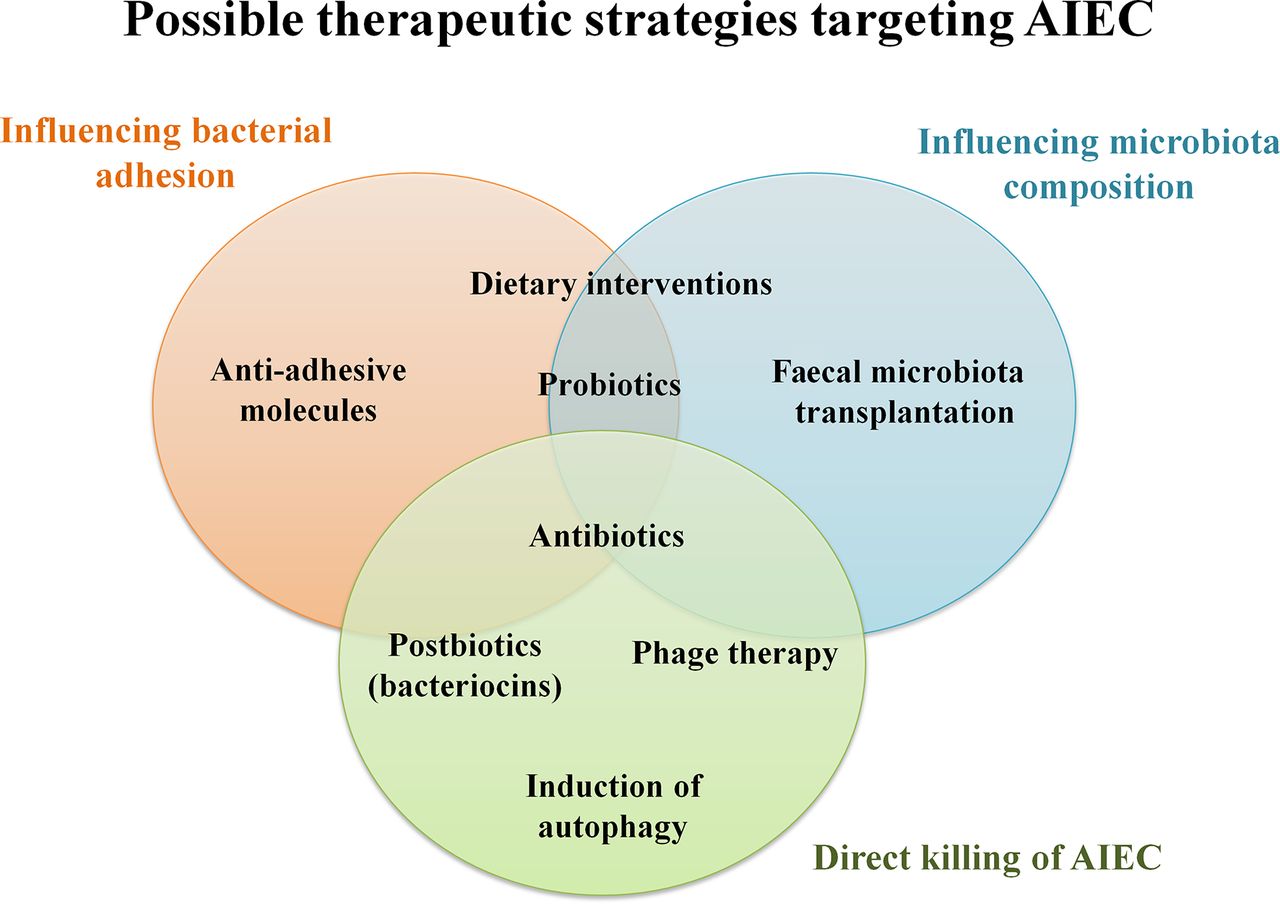
If AIEC turns out to be a disease modifier, several therapeutic strategies targeting its colonisation could potentially slow or halt the natural course of CD. An individual’s microbiota can be manipulated through nutritional interventions, antibiotic treatment, prebiotics/probiotics and postbiotics, faecal transplantation or combination therapy (figure 2). Additionally immunotherapy to restore autophagy could induce the remission of active disease or prevent relapse in the postoperative setting.175 Therapeutic interventions to modulate the microbiota after an episode of acute infectious gastroenteritis in AIEC-positive individuals may also be important to reduce the risk of CD development.155

{kind=link}
{kind=link}
Therapeutic strategies targeting adherent-invasive Escherichia coli (AIEC).
Dietary interventions
Among environmental factors, vitamin D plays a critical role in mucosal barrier homeostasis by preserving the integrity of junction complexes.176 177 In a recent study, Caco-2-bbe cells incubated with 1,25(OH)2 vitamin D3 were protected against AIEC-induced disruption of transepithelial electrical resistance and tight junction protein redistribution. Vitamin D-deficient mice treated with DSS exhibited pronounced epithelial barrier dysfunction, greater susceptibility to AIEC colonisation and exacerbated colonic injury.178 Therefore, vitamin D deficiency after AIEC infection may compromise the mucosal barrier, leading to increased susceptibility to mucosal damage and an increased risk of IBD.
Maltodextrin, a starch-derived polysaccharide and a ubiquitous dietary component, was shown to promote AIEC LF82 biofilm formation and bacterial adhesion to IECs via type 1 pili.179 Western diets are rich in polysaccharides and contain high levels of maltodextrin, which could contribute to dysbiosis and IBD susceptibility.180 Additionally, carrageenan gum (sulfated polysaccharide used as a stabiliser and bulking agent)181 and the commonly used emulsifiers polysorbate-80114 and carboxymethylcellulose,182 ha also been shown to influence bacterial adhesion, host mucosal homeostasis and gut microbiota composition and localization. This promotes the development of low-grade inflammation and favours AIEC colonisation of the gut mucosa.
Some food components like bovine lactoferrin, which is present in milk and has antimicrobial and/or anti-inflammatory properties, can inhibit bacterial adhesion and invasion.183 184 Serum-derived bovine immunoglobulin/protein isolate has recently been evoked to attenuate LF82/DSS-induced colitis and inhibit the inflammatory cascades that contribute to IBD.185 Martin et al 29 found that E coli adhesion to IEC can be inhibited with soluble plantain non-starch polysaccharide (fibre). In epidemiological studies, high fibre intake has been associated with a lower incidence of IBD and the effect of fibre on AIEC may in part explain this effect. In contrast, a high-fat and high-sugar Western diet was shown to potentiate AIEC intestinal colonisation and to promote inflammation in the CEABAC10 animal model.154 Moreover, a methyl donor-deficient diet was shown to decrease methylation levels in the CEACAM6 promoter of CEABAC10 mice, leading to abnormal CEACAM6 expression by enterocytes and increasing susceptibility to AIEC infection.186 Further investigation addressing the role of diet on AIEC intestinal colonisation is warranted.
Antibiotics
Generally, the use of traditional antibiotics has shown poor efficacy in IBD. Currently, antibiotics are indicated in patients with IBD to treat bacterial complications or in the postoperative setting of CD.187 Antibiotic therapy is less popular due to the increasing prevalence of multidrug-resistant bacteria, the exacerbation of dysbiosis and the promotion of the growth of pathogenic bacteria.188 In addition, long-term treatment with antibiotics previously used in IBD, such as metronidazole, is associated with a high risk of intolerance due to adverse effects. However, the use of antibiotics under specific conditions, such as patients colonised by AIEC strains with high disease activity, may prove to be effective. Antibiotics such as ciprofloxacin and rifaximin have fewer side effects than immunosuppressants and may be a safer alternative in patients with CD concomitantly infected with AIEC. The ongoing randomised controlled trial ‘TEOREM’ (Evaluation of Adherent Invasive E coli Eradication in Adult Crohn Disease) will assess if a 12-week treatment with ciprofloxacin and rifaximin is superior to placebo to obtain endoscopic remission in patients with ileal CD colonised with AIEC. The results from this trial can improve our understanding of whether AIEC eradication influences endoscopic remission.
Probiotics, prebiotics and postbiotics
To date, most of the clinical trials addressing the use of probiotics in CD have shown disappointing results.189 The use of probiotics that inhibit AIEC adhesion and invasion could be a potential therapeutic option to control AIEC-related gut inflammation. Boudeau et al 190 reported a significant inhibitory effect of E coli Nissle 1917 on AIEC adhesion and invasion properties. A recent study by Sivignon et al 191 showed that Saccharomyces cerevisiae CNCM I-3856 inhibited LF82 strain adhesion to the brush border of enterocytes and reduced the proinflammatory response and colitis induced in CEABAC10 mice. S cerevisiae yeasts prevent AIEC colonisation by inhibiting FimH/CEACAM6 interaction.191 Van den Abbeele et al 192 reported a reduction in AIEC survival and growth during co-culture with Lactobacillus rhamnosus GG and Lactobacillus reuteri 1063. The same authors also showed that the prebiotic long-chain arabinoxylans reduced mucin-adhesion of AIEC, while the prebiotics inulin and galacto-oligosaccharides limited AIEC growth and survival. However, isolated reports show a worsening of inflammation with probiotics in IBD mucosal explants, probably due to higher bacterial translocation.193 This highlights the need for further studies addressing the safety of these therapies in patients with IBD.
Postbiotics are soluble factors produced by probiotic bacteria and can be administered in a well-characterised manner. Bacteriocins, which are postbiotic proteins that act against a narrow spectrum of bacteria, could be an alternative therapeutic option to control AIEC. Colicins are species-specific bacteriocins that can target bacteria through different mechanisms of action. Brown et al 194 showed that colicins E1 and E9 were able to effectively kill biofilm-associated, cell-adherent and intracellular AIEC (strains LF82, HM95, 419, 615), and were superior compared with metronidazole and ciprofloxacin. Colicins do not cause damage to IEC or macrophages; however, the killing of AIEC was not associated with reduced levels of TNF-α production by infected macrophages.194 Colicins entered AIEC-containing vacuoles within macrophages by actin-mediated endocytosis mechanisms. These antibacterial agents could be delivered either as a purified protein or through colicin-producing bacteria. A recent paper reported the use of E coli Nissle 1917 as a colicin-producing probiotic.195 Recombinant replacement of a serine protease gene allowed these bacteria to secrete the selected colicin which was toxic to AIEC strains.195 However, the possible adverse effects of colicin therapy must be thoroughly studied before it is used in humans since it may result in IEC damage. E coli Nissle 1917 has been successfully used as a probiotic in a clinical trial in UC.196 Further in vivo studies on the efficacy of probiotics and their effects on AIEC are warranted. Unlike antibiotics, the selectivity of colicins suggests that they could be used to effectively eradicate a specific organism (such as AIEC), while maintaining the bacterial population that comprises the healthy gut microbiome.194
Antiadhesive molecules
The use of antiadhesive compounds against AIEC in CD is another attractive therapeutic option. Small glycomimetics molecules have been designed to saturate the carbohydrate recognition domain (CRD) of FimH by mimicking the natural ligand. These FimH antagonists block the CRD, preventing the binding of bacteria to IEC, which are washed out of the gut.197–200 One particular advantage of antiadhesive molecules is the promotion of AIEC clearance without disruption of the intestinal microbiota. The development of synthetic FimH antagonists could provide a new therapeutic approach for patients with CD who are colonised by AIEC.
Faecal microbiota transplantation
Faecal microbiota transplantation (FMT) allows modification of the microbiota composition and has been shown to be effective in recurrent C difficile infection.201 Conceptually the restoration of a ‘healthy’ intestinal microbiota could reverse the inappropriate immune mucosal stimulation in CD and create a less suitable environment for AIEC colonisation.173 The safety and efficacy of FMT in IBD are under investigation. Several independent studies have shown mixed results in IBD, probably due to differences in methodology including route, number of administrations, choice of donor and differences in disease status.202–204 Most recently, a multicentre, randomised, placebo-controlled trial showed that intensive-dosing, multidonor FMT induces clinical remission and endoscopic improvement in patients with active UC.205 To date, no investigation has addressed the impact of FMT on AIEC colonisation.
Phage therapy
Phage therapy is a biological treatment against bacterial infections, including those refractory to commonly available antibiotics. Bacteriophages are viruses that infect bacteria. Phages, unlike antibiotics, are highly specific and typically lyse a subgroup of strains within one bacterial species or within a few closely related bacterial species. Therefore, the use of phage therapy targets only a limited number of bacterial strains and may have a limited impact on the overall composition of the subject’s microbiota.206 Recent studies have demonstrated that virulent bacteriophages can replicate in gut sections from mice and are effective in reducing intestinal E coli colonisation.127 207 There is no evidence to suggest that bacteriophages are unsafe or cause serious side effects.208 A recent randomised trial of oral phage therapy in 120 children with acute bacterial diarrhoea in Bangladesh did not report any adverse events.209 Since bacteriophages are part of the human commensal microbiota and because they are highly specific, they are likely to have a better safety profile than antibiotic therapy. Galtier et al 210 found three virulent LF82 bacteriophages that were able to replicate in ileal and colon samples, and in faeces of murine specimens. A single day of oral treatment with these bacteriophages significantly decreased intestinal colonisation by LF82 AIEC strain. Promisingly, this single-dose bacteriophage cocktail reduced DSS-induced colitis symptoms over a 2-week period in mice colonised with LF82.210 Studies in human subjects are currently being planned in the USA for evaluating this treatment in patients with IBD. Namely, a phase I double-blind, randomised placebo-controlled trial is anticipated to start enrolling subjects with quiescent CD during 2017 in order to assess the safety of a lytic phage preparation containing seven bacteriophages targeting AIEC. Participants with documented AIEC in the stool will take either a phage preparation or placebo for 2 weeks to evaluate for adverse events or disease exacerbation. If safe and successful, the hope is to develop this as an adjuvant therapy in patients with CD and AIEC colonisation.
Restoration/induction of autophagy
Functional autophagy is required to restrain the intracellular replication of AIEC.71 139 140 CD is associated with polymorphisms in NOD2, IRGM and ATG16L1 genes that may render autophagy of intracellular bacteria ineffective.120 129 211 212 Lapaquette et al 71 demonstrated that the number of intramacrophagic AIEC and the secretion of proinflammatory cytokines significantly decreased after pharmacological induction of autophagy with rapamycin. These findings lead to the speculation that the stimulation of autophagy in patients with CD genotyped for risk alleles in NOD2, IRGM or ATG16L1 could be a potential treatment strategy to control the replication of AIEC and reduce the associated inflammatory response.71
Other possible therapeutic interventions
Perturbing the Enterobacteriaceae adrenergic receptor QseC with the biochemical inhibitor LED209 can attenuate experimental colitis.213 The inactivation of QseC in a pathogenic IBD-associated E coli strain can reduce its virulence in vitro and abrogate its ability to persist in a low-complexity microbiome in vivo.213 These results provide insight into the use of an antivirulence approach to target not only pathogens, but also a much larger collection of colitogenic bacteria.
Conclusion
A growing body of evidence indicates that AIEC is involved in the pathogenesis of IBD, particularly CD. Although the prevalence of AIEC in the mucosa of patients with CD varies widely across studies, several groups have reported AIEC colonisation in >50% of patients with CD. The majority of these studies have been undertaken in Europe and North America and confirmation of the presence of AIEC in patients with CD from other parts of the world will be important and are in progress. Further studies addressing the abundance of AIEC in different IBD subtypes, including CD versus UC and ileal versus colonic CD, as well as the correlation of AIEC and disease activity, will help elucidate its role in IBD and its potential use as a diagnostic tool and a therapeutic target. Therapeutic strategies directly targeting AIEC, including the use of phage therapy, bacteriocins and antiadhesive molecules, may open new avenues for the prevention and treatment of CD.
References
Footnotes
Contributors CP, CC and ZX did the bibliographic search. CP, CC, ZX, GS, JT and RH drafted the manuscript. NB, SN and JFC reviewed the manuscript. All authors contributed to the final manuscript.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.