Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression


{kind=link}
{kind=link}
{kind=link}
Abstract
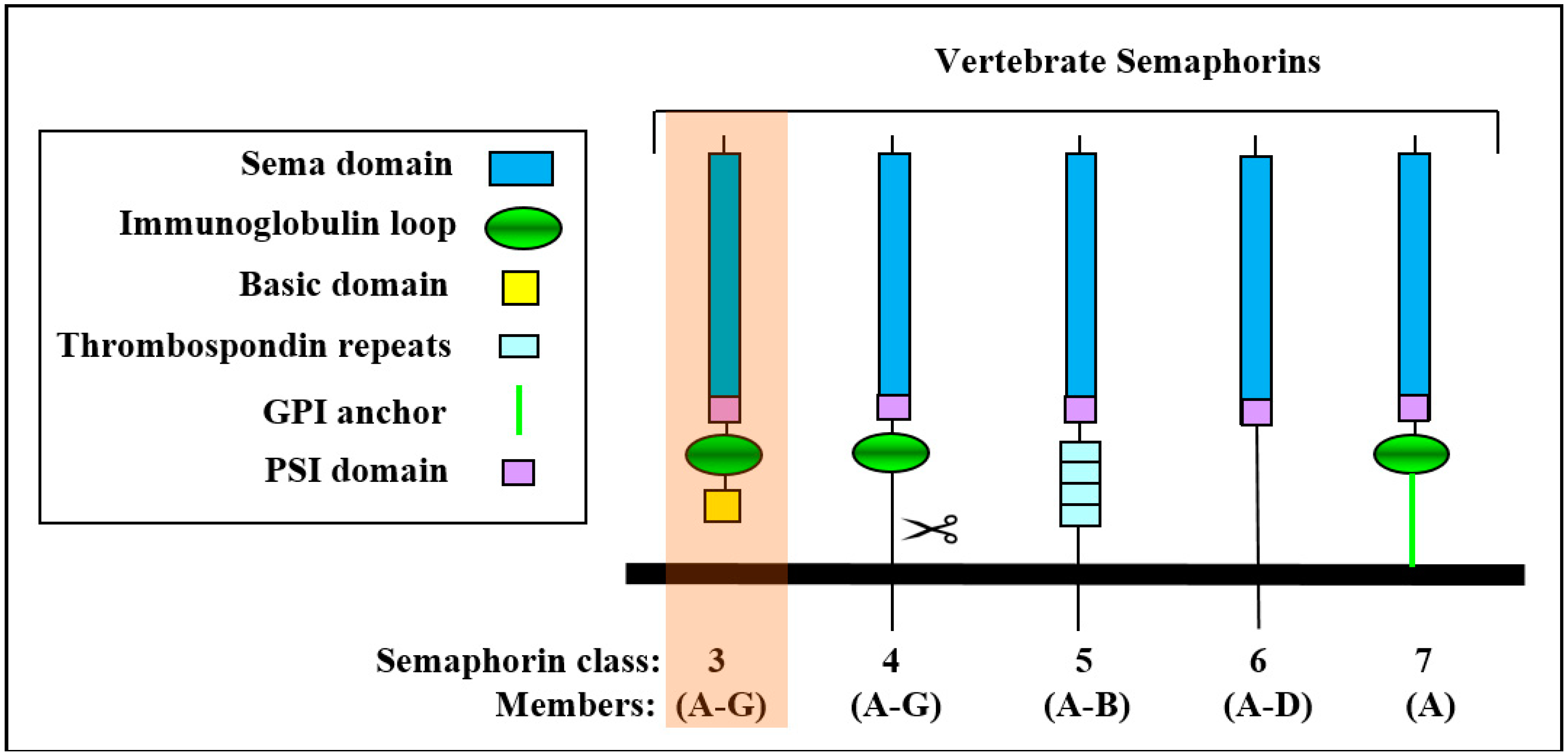
:1. The Class-3 Semaphorin Subfamily
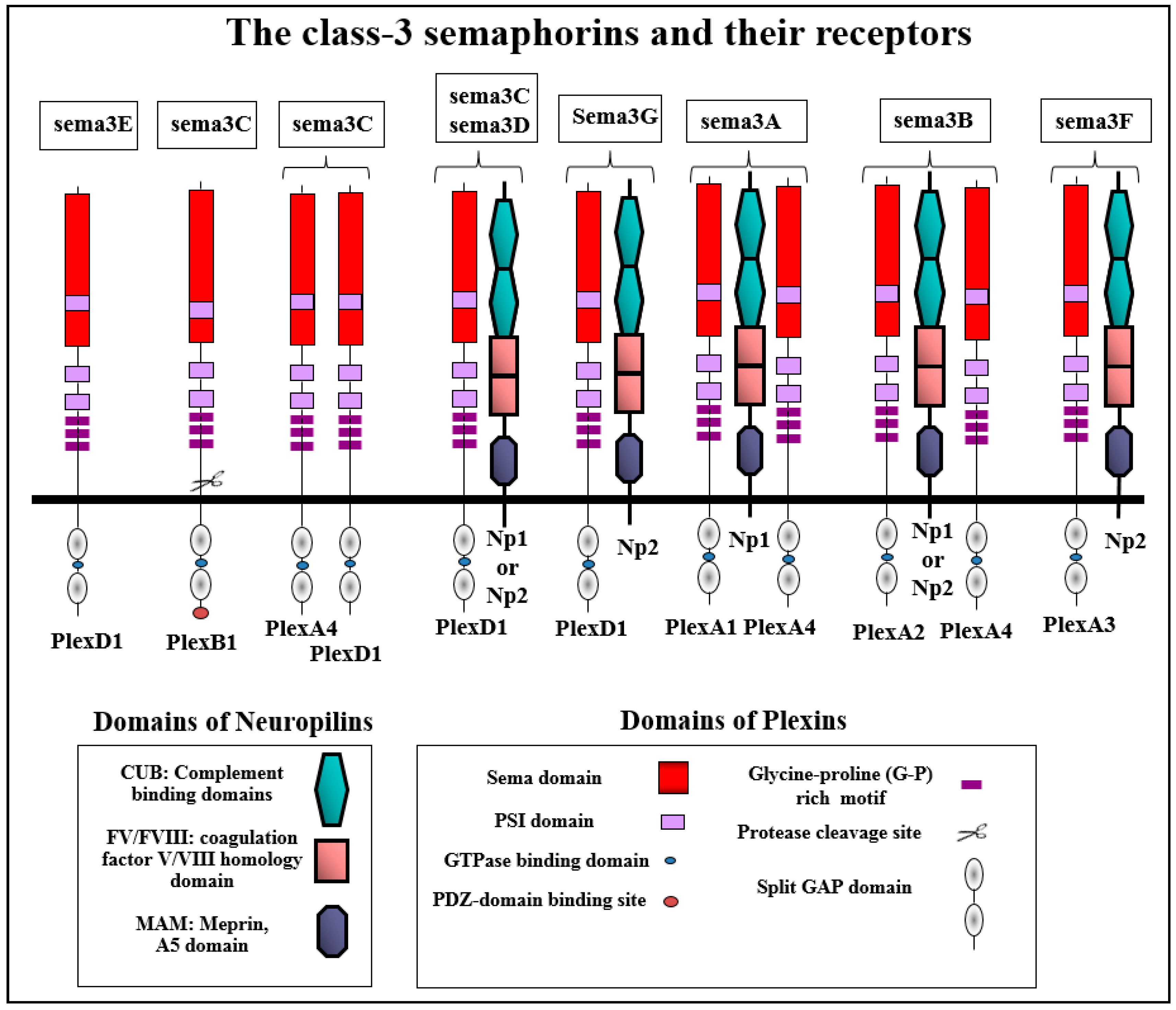
2. Class-3 Semaphorin Receptors
2.1. The Neuropilins: Multifunctional Scaffold Receptors For Class-3 Semaphorins
2.2. Plexin Receptors: Signal Transducers of Class-3 Semaphorins
3. Class-3 Semaphorins as Regulators of Tumor Progression
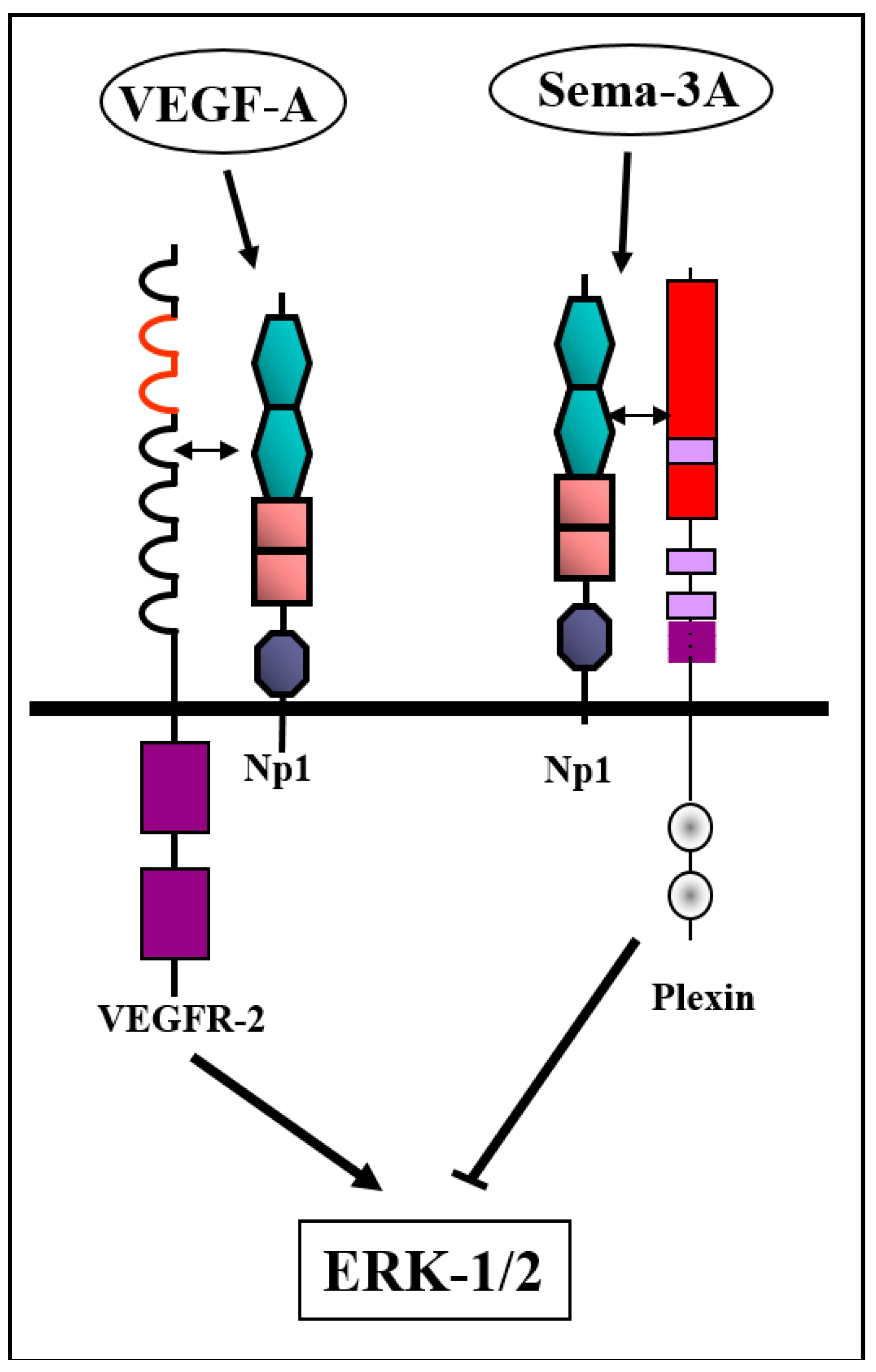
3.1. Modulation of Tumor Angiogenesis by Class-3 Semaphorins
3.2. Effects of Class-3 Semaphorins on Lymphangiogenesis
3.3. Modulation of Tumor Progression by Direct Effects on Tumor Cells
3.4. Modulation of Tumor Progression by Mechanisms Involving Stromal Cells Other Than Endothelial Cells
3.5. Modulation of Tumor Progression by Class-3 Semaphorins that Modulate Immune Responses
4. Modifications of Class-3 Semaphorins and Class-3 Semaphorin Receptor Genes As Modulators of Tumor Progression
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Neufeld, G.; Kessler, O. The semaphorins: Versatile regulators of tumour progression and tumour angiogenesis. Nat. Rev. Cancer 2008, 8, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Feiner, L.; Koppel, A.M.; Kobayashi, H.; Raper, J.A. Secreted chick semaphorins bind recombinant neuropilin with similar affinities but bind different subsets of neurons in situ. Neuron 1997, 19, 539–545. [Google Scholar] [CrossRef]
- Love, C.A.; Harlos, K.; Mavaddat, N.; Davis, S.J.; Stuart, D.I.; Jones, E.Y.; Esnouf, R.M. The ligand-binding face of the semaphorins revealed by the high-resolution crystal structure of SEMA4D. Nat. Struct. Biol. 2003, 10, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Antipenko, A.; Himanen, J.P.; van Leyen, K.; Nardi-Dei, V.; Lesniak, J.; Barton, W.A.; Rajashankar, K.R.; Lu, M.; Hoemme, C.; Puschel, A.W.; et al. Structure of the Semaphorin-3A Receptor Binding Module. Neuron 2003, 39, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Juo, Z.S.; Shim, A.H.; Focia, P.J.; Chen, X.; Garcia, K.C.; He, X. Structural Basis of Semaphorin-Plexin Recognition and Viral Mimicry from Sema7A and A39R Complexes with PlexinC1. Cell 2010, 142, 749–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppel, A.M.; Raper, J.A. Collapsin-1 covalently dimerizes, and dimerization is necessary for collapsing activity. J. Biol. Chem. 1998, 273, 15708–15713. [Google Scholar] [CrossRef] [PubMed]
- Janssen, B.J.; Malinauskas, T.; Weir, G.A.; Cader, M.Z.; Siebold, C.; Jones, E.Y. Neuropilins lock secreted semaphorins onto plexins in a ternary signaling complex. Nat. Struct. Mol. Biol. 2012, 19, 1293–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klostermann, A.; Lohrum, M.; Adams, R.H.; Puschel, A.W. The chemorepulsive activity of the axonal guidance signal semaphorin D requires dimerization. J. Biol. Chem. 1998, 273, 7326–7331. [Google Scholar] [CrossRef]
- Nogi, T.; Yasui, N.; Mihara, E.; Matsunaga, Y.; Noda, M.; Yamashita, N.; Toyofuku, T.; Uchiyama, S.; Goshima, Y.; Kumanogoh, A.; et al. Structural basis for semaphorin signalling through the plexin receptor. Nature 2010, 467, 1123–1127. [Google Scholar] [CrossRef]
- Adams, R.H.; Lohrum, M.; Klostermann, A.; Betz, H.; Puschel, A.W. The chemorepulsive activity of secreted semaphorins is regulated by furin-dependent proteolytic processing. EMBO J. 1997, 16, 6077–6086. [Google Scholar] [CrossRef] [Green Version]
- Varshavsky, A.; Kessler, O.; Abramovitch, S.; Kigel, B.; Zaffryar, S.; Akiri, G.; Neufeld, G. Semaphorin-3B Is an Angiogenesis Inhibitor That Is Inactivated by Furin-Like Pro-Protein Convertases. Cancer Res. 2008, 68, 6922–6931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, C.; Ambartsumian, N.; Gilestro, G.; Thomsen, B.; Comoglio, P.; Tamagnone, L.; Guldberg, P.; Lukanidin, E. Proteolytic Processing Converts the Repelling Signal Sema3E into an Inducer of Invasive Growth and Lung Metastasis. Cancer Res. 2005, 65, 6167–6177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casazza, A.; Kigel, B.; Maione, F.; Capparuccia, L.; Kessler, O.; Giraudo, E.; Mazzone, M.; Neufeld, G.; Tamagnone, L. Tumour growth inhibition and anti-metastatic activity of a mutated furin-resistant Semaphorin 3E isoform. EMBO Mol. Med. 2012, 4, 234–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casazza, A.; Finisguerra, V.; Capparuccia, L.; Camperi, A.; Swiercz, J.M.; Rizzolio, S.; Rolny, C.; Christensen, C.; Bertotti, A.; Sarotto, I.; et al. Sema3E-Plexin D1 signaling drives human cancer cell invasiveness and metastatic spreading in mice. J. Clin. Investig. 2010, 120, 2684–2698. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.W.; Hellman, L.M.; Xu, P.; Fried, M.G.; Vander Kooi, C.W. Furin Processing of Semaphorin 3F Determines its Anti-Angiogenic Activity by Regulating Direct Binding and Competition for Neuropilin. Biochemistry 2010, 18, 4068–4075. [Google Scholar] [CrossRef] [PubMed]
- Valiulyte, I.; Preitakaite, V.; Tamasauskas, A.; Kazlauskas, A. Importance of the putative furin recognition site 742RNRR745 for antiangiogenic Sema3C activity in vitro. Braz. J. Med. Biol. Res. 2018, 51, e7786. [Google Scholar] [CrossRef]
- Tamagnone, L.; Artigiani, S.; Chen, H.; He, Z.; Ming, G.I.; Song, H.; Chedotal, A.; Winberg, M.L.; Goodman, C.S.; Poo, M.; et al. Plexins are a large family of receptors for transmembrane, secreted, and GPI-anchored semaphorins in vertebrates. Cell 1999, 99, 71–80. [Google Scholar] [CrossRef]
- Gu, C.; Yoshida, Y.; Livet, J.; Reimert, D.V.; Mann, F.; Merte, J.; Henderson, C.E.; Jessell, T.M.; Kolodkin, A.L.; Ginty, D.D. Semaphorin 3E and plexin-D1 control vascular pattern independently of neuropilins. Science 2005, 307, 265–268. [Google Scholar] [CrossRef]
- He, Z.; Tessier-Lavigne, M. Neuropilin is a receptor for the axonal chemorepellent Semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef]
- Chen, H.; Chedotal, A.; He, Z.; Goodman, C.S.; Tessier-Lavigne, M. Neuropilin-2, a novel member of the neuropilin family, is a high affinity receptor for the semaphorins Sema E and Sema IV but not Sema III. Neuron 1997, 19, 547–559. [Google Scholar] [CrossRef]
- Takahashi, T.; Fournier, A.; Nakamura, F.; Wang, L.H.; Murakami, Y.; Kalb, R.G.; Fujisawa, H.; Strittmatter, S.M. Plexin-neuropilin-1 complexes form functional semaphorin-3A receptors. Cell 1999, 99, 59–69. [Google Scholar] [CrossRef]
- Kolodkin, A.L.; Levengood, D.V.; Rowe, E.G.; Tai, Y.T.; Giger, R.J.; Ginty, D.D. Neuropilin is a semaphorin III receptor. Cell 1997, 90, 753–762. [Google Scholar] [CrossRef]
- Smolkin, T.; Nir-Zvi, I.; Duvshani, N.; Mumblat, Y.; Kessler, O.; Neufeld, G. Complexes of plexin-A4 and plexin-D1 convey semaphorin-3C signals to induce cytoskeletal collapse in the absence of neuropilins. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed]
- Gitler, A.D.; Lu, M.M.; Epstein, J.A. PlexinD1 and semaphorin signaling are required in endothelial cells for cardiovascular development. Dev. Cell 2004, 7, 107–116. [Google Scholar] [CrossRef]
- Gitay-Goren, H.; Cohen, T.; Tessler, S.; Soker, S.; Gengrinovitch, S.; Rockwell, P.; Klagsbrun, M.; Levi, B.-Z.; Neufeld, G. Selective binding of VEGF121 to one of the three VEGF receptors of vascular endothelial cells. J. Biol. Chem. 1996, 271, 5519–5523. [Google Scholar] [CrossRef] [PubMed]
- Soker, S.; Takashima, S.; Miao, H.Q.; Neufeld, G.; Klagsbrun, M. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform specific receptor for vascular endothelial growth factor. Cell 1998, 92, 735–745. [Google Scholar] [CrossRef]
- Gluzman-Poltorak, Z.; Cohen, T.; Herzog, Y.; Neufeld, G. Neuropilin-2 and Neuropilin-1 are receptors for 165-amino acid long form of vascular endothelial growth factor (VEGF) and of placenta growth factor-2, but only neuropilin-2 functions as a receptor for the 145 amino acid form of VEGF. J. Biol. Chem. 2000, 275, 18040–18045. [Google Scholar] [CrossRef] [PubMed]
- Makinen, T.; Olofsson, B.; Karpanen, T.; Hellman, U.; Soker, S.; Klagsbrun, M.; Eriksson, U.; Alitalo, K. Differential binding of vascular endothelial growth factor B splice and proteolytic isoforms to neuropilin-1. J. Biol. Chem. 1999, 274, 21217–21222. [Google Scholar] [CrossRef] [PubMed]
- Karpanen, T.; Heckman, C.A.; Keskitalo, S.; Jeltsch, M.; Ollila, H.; Neufeld, G.; Tamagnone, L.; Alitalo, K. Functional interaction of VEGF-C and VEGF-D with neuropilin receptors. FASEB J. 2006, 20, 1462–1472. [Google Scholar] [CrossRef] [PubMed]
- Migdal, M.; Huppertz, B.; Tessler, S.; Comforti, A.; Shibuya, M.; Reich, R.; Baumann, H.; Neufeld, G. Neuropilin-1 is a placenta growth factor-2 receptor. J. Biol. Chem. 1998, 273, 22272–22278. [Google Scholar] [CrossRef] [PubMed]
- Sulpice, E.; Plouet, J.; Berge, M.; Allanic, D.; Tobelem, G.; Merkulova-Rainon, T. Neuropilin-1 and neuropilin-2 act as coreceptors, potentiating proangiogenic activity. Blood 2007, 111, 2036–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhl, L.; Folestad, E.B.; Gladh, H.; Wang, Y.; Moessinger, C.; Jakobsson, L.; Eriksson, U. Neuropilin 1 binds platelet-derived growth factor (PDGF)-D and is a co-receptor in PDGF-D/PDGF receptor beta signaling. J. Cell Sci. 2017, 130, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Glinka, Y.; Stoilova, S.; Mohammed, N.; Prud’homme, G.J. Neuropilin-1 exerts coreceptor function for TGF-beta-1 on the membrane of cancer cells and enhances responses to both latent and active TGF-beta. Carcinogenesis 2011, 32, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.H.; Ying, N.W.; Wu, M.H.; Chiang, W.F.; Hsu, C.L.; Wong, T.Y.; Jin, Y.T.; Hong, T.M.; Chen, Y.L. Galectin-1, a novel ligand of neuropilin-1, activates VEGFR-2 signaling and modulates the migration of vascular endothelial cells. Oncogene 2008, 27, 3746–3753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluzman-Poltorak, Z.; Cohen, T.; Shibuya, M.; Neufeld, G. Vascular endothelial growth factor receptor-1 and neuropilin-2 form complexes. J. Biol. Chem. 2001, 276, 18688–18694. [Google Scholar] [CrossRef] [PubMed]
- Favier, B.; Alam, A.; Barron, P.; Bonnin, J.; Laboudie, P.; Fons, P.; Mandron, M.; Herault, J.P.; Neufeld, G.; Savi, P.; et al. Neuropilin-2 interacts with VEGFR-2 and VEGFR-3 and promotes human endothelial cell survival and migration. Blood 2006, 108, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsaka, A.; Hirota-Komatsu, S.; Araki, M.; Komatsu, N. Platelet-derived growth factor receptors form complexes with neuropilin-1 during megakaryocytic differentiation of thrombopoietin-dependent UT-7/TPO cells. Biochem. Biophys. Res. Commun. 2015, 459, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Bayley, C.; Shuttleworth, C.A.; Kielty, C.M. Neuropilin-1 regulates platelet-derived growth factor receptor signaling in mesenchymal stem cells. Biochem. J. 2010, 427, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Ben-Zvi, A.; Ben-Gigi, L.; Klein, H.; Behar, O. Modulation of semaphorin3A activity by p75 neurotrophin receptor influences peripheral axon patterning. J. Neurosci. 2007, 27, 13000–13011. [Google Scholar] [CrossRef]
- Fukasawa, M.; Matsushita, A.; Korc, M. Neuropilin-1 Interacts with Integrin beta1 and Modulates Pancreatic Cancer Cell Growth, Survival and Invasion. Cancer Biol. Ther. 2007, 6, 1173–1180. [Google Scholar] [CrossRef]
- Robinson, S.D.; Reynolds, L.E.; Kostourou, V.; Reynolds, A.R.; da Silva, R.G.; Tavora, B.; Baker, M.; Marshall, J.F.; Hodivala-Dilke, K.M. Alphavbeta3-integrin limits the contribution of neuropilin-1 to VEGF-induced angiogenesis. J. Biol. Chem. 2009, 284, 33966–33981. [Google Scholar] [CrossRef] [PubMed]
- Castellani, V.; De Angelis, E.; Kenwrick, S.; Rougon, G. Cis and trans interactions of L1 with neuropilin-1 control axonal responses to semaphorin 3A. EMBO J. 2002, 21, 6348–6357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, A.G.; Demyanenko, G.P.; Powell, A.; Schachner, M.; Enriquez-Barreto, L.; Tran, T.S.; Polleux, F.; Maness, P.F. Close homolog of L1 and neuropilin 1 mediate guidance of thalamocortical axons at the ventral telencephalon. J. Neurosci. 2007, 27, 13667–13679. [Google Scholar] [CrossRef]
- Falk, J.; Bechara, A.; Fiore, R.; Nawabi, H.; Zhou, H.; Hoyo-Becerra, C.; Bozon, M.; Rougon, G.; Grumet, M.; Puschel, A.W.; et al. Dual Functional Activity of Semaphorin 3B Is Required for Positioning the Anterior Commissure. Neuron 2005, 48, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhau, H.E.; Osunkoya, A.O.; Iqbal, S.; Yang, X.; Fan, S.; Chen, Z.; Wang, R.; Marshall, F.F.; Chung, L.W.; et al. Vascular endothelial growth factor regulates myeloid cell leukemia-1 expression through neuropilin-1-dependent activation of c-MET signaling in human prostate cancer cells. Mol. Cancer 2010, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appleton, B.A.; Wu, P.; Maloney, J.; Yin, J.; Liang, W.C.; Stawicki, S.; Mortara, K.; Bowman, K.K.; Elliott, J.M.; Desmarais, W.; et al. Structural studies of neuropilin/antibody complexes provide insights into semaphorin and VEGF binding. EMBO J. 2007, 26, 4902–4912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttmann-Raviv, N.; Shraga-Heled, N.; Varshavsky, A.; Guimaraes-Sternberg, C.; Kessler, O.; Neufeld, G. Semaphorin-3A and Semaphorin-3F Work Together to Repel Endothelial Cells and to Inhibit Their Survival by Induction of Apoptosis. J. Biol. Chem. 2007, 282, 26294–26305. [Google Scholar] [CrossRef] [Green Version]
- Mumblat, Y.; Kessler, O.; Ilan, N.; Neufeld, G. Full length semaphorin-3C functions as an inhibitor of tumor lymphangiogenesis and tumor metastasis. Cancer Res. 2015, 75, 2177–2186. [Google Scholar] [CrossRef]
- Guo, H.F.; Li, X.; Parker, M.W.; Waltenberger, J.; Becker, P.M.; Vander Kooi, C.W. Mechanistic Basis for the Potent Anti-angiogenic Activity of Semaphorin 3F. Biochemistry 2013, 52, 7551–7558. [Google Scholar] [CrossRef]
- Lanahan, A.; Zhang, X.; Fantin, A.; Zhuang, Z.; Rivera-Molina, F.; Speichinger, K.; Prahst, C.; Zhang, J.; Wang, Y.; Davis, G.; et al. The Neuropilin 1 Cytoplasmic Domain Is Required for VEGF-A-Dependent Arteriogenesis. Dev. Cell 2013, 25, 156–168. [Google Scholar] [CrossRef] [Green Version]
- Yaqoob, U.; Cao, S.; Shergill, U.; Jagavelu, K.; Geng, Z.; Yin, M.; de Assuncao, T.M.; Cao, Y.; Szabolcs, A.; Thorgeirsson, S.; et al. Neuropilin-1 stimulates tumor growth by increasing fibronectin fibril assembly in the tumor microenvironment. Cancer Res. 2012, 72, 4047–4059. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.B.; Reed, R.R. Cloning and characterization of neuropilin-1-interacting protein: A PSD-95/Dlg/ZO-1 domain-containing protein that interacts with the cytoplasmic domain of neuropilin-1. J. Neurosci. 1999, 19, 6519–6527. [Google Scholar] [CrossRef] [PubMed]
- Prahst, C.; Heroult, M.; Lanahan, A.A.; Uziel, N.; Kessler, O.; Shraga-Heled, N.; Simons, M.; Neufeld, G.; Augustin, H.G. Neuropilin-1/VEGFR-2 complexing requires the PDZ-binding domain of neuropilin-1. J. Biol. Chem. 2008, 283, 25110–25114. [Google Scholar] [CrossRef] [PubMed]
- Hota, P.K.; Buck, M. Plexin structures are coming: Opportunities for multilevel investigations of semaphorin guidance receptors, their cell signaling mechanisms, and functions. Cell. Mol. Life Sci. 2012, 69, 3765–3805. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Strittmatter, S.M. PlexinA1 Autoinhibition by the Plexin Sema Domain. Neuron 2001, 29, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Oinuma, I.; Ishikawa, Y.; Katoh, H.; Negishi, M. The Semaphorin 4D receptor Plexin-B1 is a GTPase activating protein for R-Ras. Science 2004, 305, 862–865. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yang, T.; Terman, J.R.; Zhang, X. Crystal structure of the plexin A3 intracellular region reveals an autoinhibited conformation through active site sequestration. Proc. Natl. Acad. Sci. USA 2009, 106, 15610–15615. [Google Scholar] [CrossRef]
- Sakurai, A.; Gavard, J.; nnas-Linhares, Y.; Basile, J.R.; Amornphimoltham, P.; Palmby, T.R.; Yagi, H.; Zhang, F.; Randazzo, P.A.; Li, X.; et al. Semaphorin 3E initiates anti-angiogenic signaling through Plexin-D1 by regulating Arf6 and R-Ras. Mol. Cell. Biol. 2010, 30, 3086–3098. [Google Scholar] [CrossRef]
- Worzfeld, T.; Swiercz, J.M.; Senturk, A.; Genz, B.; Korostylev, A.; Deng, S.; Xia, J.; Hoshino, M.; Epstein, J.A.; Chan, A.M.; et al. Genetic dissection of plexin signaling in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 2194–2199. [Google Scholar] [CrossRef] [Green Version]
- Janssen, B.J.; Robinson, R.A.; Perez-Branguli, F.; Bell, C.H.; Mitchell, K.J.; Siebold, C.; Jones, E.Y. Structural basis of semaphorin-plexin signalling. Nature 2010, 467, 1118–1122. [Google Scholar] [CrossRef]
- Kigel, B.; Rabinowicz, N.; Varshavsky, A.; Kessler, O.; Neufeld, G. Plexin-A4 promotes tumor progression and tumor angiogenesis by enhancement of VEGF and bFGF signaling. Blood 2011, 118, 4285–4296. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, Q.; Waimey, K.E.; Golding, M.; Takamatsu, H.; Kumanogoh, A.; Fujisawa, H.; Cheng, H.J.; Ruhrberg, C. Plexin A3 and plexin A4 convey semaphorin signals during facial nerve development. Dev. Biol. 2008, 324, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, H.; Lei, Y.; Eun, S.-Y.; Ting, J.P. Plexin-A4 semaphorin 3A signaling is required for Toll-like receptor- and sepsis-induced cytokine storm. J. Exp. Med. 2010, 207, 2943–2957. [Google Scholar] [CrossRef] [PubMed]
- Bouvree, K.; Brunet, I.; Del, T.R.; Gordon, E.; Prahst, C.; Cristofaro, B.; Mathivet, T.; Xu, Y.; Soueid, J.; Fortuna, V.; et al. Semaphorin3A, Neuropilin-1, and PlexinA1 are Required for Lymphatic Valve Formation. Circ. Res. 2012, 111, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Moretti, S.; Procopio, A.; Lazzarini, R.; Rippo, M.R.; Testa, R.; Marra, M.; Tamagnone, L.; Catalano, A. Semaphorin3A signaling controls Fas (CD95)-mediated apoptosis by promoting Fas translocation into lipid rafts. Blood 2007, 111, 2290–2299. [Google Scholar] [CrossRef] [Green Version]
- Sabag, A.D.; Smolkin, T.; Mumblat, Y.; Ueffing, M.; Kessler, O.; Gloeckner, C.J.; Neufeld, G. The role of the plexin-A2 receptor in Sema3A and Sema3B signal transduction. J. Cell Sci. 2014, 127, 5240–5252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Zvi, A.; Manor, O.; Schachner, M.; Yaron, A.; Tessier-Lavigne, M.; Behar, O. The Semaphorin Receptor PlexinA3 Mediates Neuronal Apoptosis during Dorsal Root Ganglia Development. J. Neurosci. 2008, 28, 12427–12432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Uemura, A.; Fukushima, Y.; Yoshida, Y.; Hirashima, M. Semaphorin 3G Provides a Repulsive Guidance Cue to Lymphatic Endothelial Cells via Neuropilin-2/PlexinD1. Cell Rep. 2016, 17, 2299–2311. [Google Scholar] [CrossRef] [Green Version]
- Hamm, M.J.; Kirchmaier, B.C.; Herzog, W. Sema3d controls collective endothelial cell migration by distinct mechanisms via Nrp1 and PlxnD1. J. Cell Biol. 2016, 215, 415–430. [Google Scholar] [CrossRef] [Green Version]
- Aghajanian, H.; Choi, C.; Ho, V.C.; Gupta, M.; Singh, M.K.; Epstein, J.A. Sema3D and Sema3E direct endothelial motility through distinct molecular signaling pathways. J. Biol. Chem. 2014, 289, 17971–17979. [Google Scholar] [CrossRef]
- Peacock, J.W.; Takeuchi, A.; Hayashi, N.; Liu, L.; Tam, K.J.; Al, N.N.; Khazamipour, N.; Tombe, T.; Dejima, T.; Lee, K.C.; et al. SEMA3C drives cancer growth by transactivating multiple receptor tyrosine kinases via Plexin B1. EMBO Mol. Med. 2018, 10, 219–238. [Google Scholar] [CrossRef] [Green Version]
- Negishi, M.; Oinuma, I.; Katoh, H. R-ras as a key player for signaling pathway of plexins. Mol. Neurobiol. 2005, 32, 217–222. [Google Scholar] [CrossRef]
- Toyofuku, T.; Yoshida, J.; Sugimoto, T.; Zhang, H.; Kumanogoh, A.; Hori, M.; Kikutani, H. FARP2 triggers signals for Sema3A-mediated axonal repulsion. Nat. Neurosci. 2005, 8, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, A.; Jian, X.; Lee, C.J.; Manavski, Y.; Chavakis, E.; Donaldson, J.; Randazzo, P.A.; Gutkind, J.S. Phosphatidylinositol-4-phosphate 5-Kinase and GEP100/Brag2 Protein Mediate Antiangiogenic Signaling by Semaphorin 3E-Plexin-D1 through Arf6 Protein. J. Biol. Chem. 2011, 286, 34335–34345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terman, J.R.; Mao, T.; Pasterkamp, R.J.; Yu, H.H.; Kolodkin, A.L. MICALs, a family of conserved flavoprotein oxidoreductases, function in plexin-mediated axonal repulsion. Cell 2002, 109, 887–900. [Google Scholar] [CrossRef]
- Hung, R.J.; Yazdani, U.; Yoon, J.; Wu, H.; Yang, T.; Gupta, N.; Huang, Z.; van Berkel, W.J.; Terman, J.R. Mical links semaphorins to F-actin disassembly. Nature 2010, 463, 823–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, R.J.; Pak, C.W.; Terman, J.R. Direct Redox Regulation of F-Actin Assembly and Disassembly by Mical. Science 2011, 334, 1710–1713. [Google Scholar] [CrossRef]
- Hung, R.J.; Spaeth, C.S.; Yesilyurt, H.G.; Terman, J.R. SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat. Cell Biol. 2013, 15, 1445–1454. [Google Scholar] [CrossRef] [Green Version]
- Franco, M.; Tamagnone, L. Tyrosine phosphorylation in semaphorin signalling: Shifting into overdrive. EMBO Rep. 2008, 9, 865–871. [Google Scholar] [CrossRef]
- Puschel, A.W. GTPases in semaphorin signaling. Adv. Exp. Med. Biol. 2007, 600, 12–23. [Google Scholar]
- Folkman, J. What is the evidence that tumors are angiogenesis dependent. J. Nat. Cancer Inst. 1990, 82, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Kessler, O.; Shraga-Heled, N.; Lange, T.; Gutmann-Raviv, N.; Sabo, E.; Baruch, L.; Machluf, M.; Neufeld, G. Semaphorin-3F Is an Inhibitor of Tumor Angiogenesis. Cancer Res. 2004, 64, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Bielenberg, D.R.; Hida, Y.; Shimizu, A.; Kaipainen, A.; Kreuter, M.; Kim, C.C.; Klagsbrun, M. Semaphorin 3F, a chemorepulsant for endothelial cells, induces a poorly vascularized, encapsulated, nonmetastatic tumor phenotype. J. Clin. Investig. 2004, 114, 1260–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, L.M.; Barillas, S.; Weis, S.M.; Gothert, J.R.; Cheresh, D.A. Semaphorin 3A suppresses VEGF-mediated angiogenesis yet acts as a vascular permeability factor. Blood 2008, 111, 2674–2680. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.J.; Hu, J.; Uemura, A.; Tetzlaff, F.; Augustin, H.G.; Fischer, A. Semaphorin-3C signals through Neuropilin-1 and PlexinD1 receptors to inhibit pathological angiogenesis. EMBO Mol. Med. 2015, 20, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Sabag, A.D.; Bode, J.; Fink, D.; Kigel, B.; Kugler, W.; Neufeld, G. Semaphorin-3D and Semaphorin-3E Inhibit the Development of Tumors from Glioblastoma Cells Implanted in the Cortex of the Brain. PLoS ONE 2012, 7, e42912. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Folkman, J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef]
- Maione, F.; Molla, F.; Meda, C.; Latini, R.; Zentilin, L.; Giacca, M.; Seano, G.; Serini, G.; Bussolino, F.; Giraudo, E. Semaphorin 3A is an endogenous angiogenesis inhibitor that blocks tumor growth and normalizes tumor vasculature in transgenic mouse models. J Clin. Investig. 2009, 119, 3356–3372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gioelli, N.; Maione, F.; Camillo, C.; Ghitti, M.; Valdembri, D.; Morello, N.; Darche, M.; Zentilin, L.; Cagnoni, G.; Qiu, Y.; et al. A rationally designed NRP1-independent superagonist SEMA3A mutant is an effective anticancer agent. Sci. Transl. Med. 2018, 10, 10–442. [Google Scholar] [CrossRef] [PubMed]
- Vacca, A.; Scavelli, C.; Serini, G.; Di, P.G.; Cirulli, T.; Merchionne, F.; Ribatti, D.; Bussolino, F.; Guidolin, D.; Piaggio, G.; et al. Loss of inhibitory semaphorin 3A (SEMA3A) autocrine loops in bone marrow endothelial cells of patients with multiple myeloma. Blood 2006, 108, 1661–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavi, N.; Kessler, O.; Ziv, K.; Nir-Zvi, I.; Mumblat, Y.; Eiza, N.; Paran, Y.; Brenner, B.; Vadasz, Z.; Neufeld, G. Semaphorin-3A inhibits multiple myeloma progression in a mouse model. Carcinogenesis 2018, 39, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.G.; Wen, R.T.; Qi, K.; Li, J.; Zheng, G.X.; Wang, Y.F.; Hong, Y.G.; Zhang, Y.M. The neuropilin-1 ligand, Sema3A, acts as a tumor suppressor in the pathogenesis of acute leukemia. Anat. Rec. (Hoboken) 2018. [Google Scholar] [CrossRef] [PubMed]
- Toledano, S.; Lu, H.; Palacio, A.; Kigel, B.; Kessler, O.; Allon, G.; Barak, Y.; Neufeld, G.; Schaal, S. A SEMA3E mutant resistant to cleavage by furins (UNCL-SEMA3E) inhibits choroidal neovascularization. Exp. Eye Res. 2016, 153, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Oh, W.J.; Gaiano, N.; Yoshida, Y.; Gu, C. Semaphorin 3E-Plexin-D1 signaling regulates VEGF function in developmental angiogenesis via a feedback mechanism. Genes Dev. 2011, 25, 1399–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zygmunt, T.; Gay, C.M.; Blondelle, J.; Singh, M.K.; Flaherty, K.M.; Means, P.C.; Herwig, L.; Krudewig, A.; Belting, H.G.; Affolter, M.; et al. Semaphorin-PlexinD1 Signaling Limits Angiogenic Potential via the VEGF Decoy Receptor sFlt1. Dev. Cell 2011, 21, 301–314. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Limberg, B.J.; Whitaker, G.B.; Perman, B.; Leahy, D.J.; Rosenbaum, J.S.; Ginty, D.D.; Kolodkin, A.L. Characterization of neuropilin-1 structural features that confer binding to semaphorin 3A and vascular endothelial growth factor 165. J. Biol. Chem. 2002, 277, 18069–18076. [Google Scholar] [CrossRef]
- Meadows, S.M.; Fletcher, P.J.; Moran, C.; Xu, K.; Neufeld, G.; Chauvet, S.; Mann, F.; Krieg, P.A.; Cleaver, O. Integration of repulsive guidance cues generates avascular zones that shape Mammalian blood vessels. Circ. Res. 2012, 110, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Xiang, R.H.; Hensel, C.H.; Garcia, D.K.; Carlson, H.C.; Kok, K.; Daly, M.C.; Kerbacher, K.; van den, B.A.; Veldhuis, P.; Buys, C.H.; et al. Isolation of the human semaphorin III/F gene (SEMA3F) at chromosome 3p21, a region deleted in lung cancer. Genomics 1996, 32, 39–48. [Google Scholar] [CrossRef]
- Tomizawa, Y.; Sekido, Y.; Kondo, M.; Gao, B.; Yokota, J.; Roche, J.; Drabkin, H.; Lerman, M.I.; Gazdar, A.F.; Minna, J.D. Inhibition of lung cancer cell growth and induction of apoptosis after reexpression of 3p21.3 candidate tumor suppressor gene SEMA3B. Proc. Natl. Acad. Sci. USA 2001, 98, 13954–13959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beuten, J.; Garcia, D.; Brand, T.C.; He, X.; Balic, I.; Canby-Hagino, E.; Troyer, D.A.; Baillargeon, J.; Hernandez, J.; Thompson, I.M.; et al. Semaphorin 3B and 3F Single Nucleotide Polymorphisms are Associated With Prostate Cancer Risk and Poor Prognosis. J. Urol. 2009, 182, 1614–1620. [Google Scholar] [CrossRef] [PubMed]
- Kusy, S.; Potiron, V.; Zeng, C.; Franklin, W.; Brambilla, E.; Minna, J.; Drabkin, H.A.; Roche, J. Promoter characterization of Semaphorin SEMA3F, a tumor suppressor gene. Biochim. Biophys. Acta 2005, 1730, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Futamura, M.; Kamino, H.; Miyamoto, Y.; Kitamura, N.; Nakamura, Y.; Ohnishi, S.; Masuda, Y.; Arakawa, H. Possible Role of Semaphorin 3F, a Candidate Tumor Suppressor Gene at 3p21.3, in p53-Regulated Tumor Angiogenesis Suppression. Cancer Res. 2007, 67, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Shirvan, A.; Ziv, I.; Fleminger, G.; Shina, R.; He, Z.G.; Brudo, I.; Melamed, E.; Barzilai, A. Semaphorins as mediators of neuronal apoptosis. J. Neurochem. 1999, 73, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Reidy, K.J.; Villegas, G.; Teichman, J.; Veron, D.; Shen, W.; Jimenez, J.; Thomas, D.; Tufro, A. Semaphorin3a regulates endothelial cell number and podocyte differentiation during glomerular development. Development 2009, 136, 3979–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagnard, D.; Vaillant, C.; Khuth, S.T.; Dufay, N.; Lohrum, M.; Puschel, A.W.; Belin, M.F.; Bolz, J.; Thomasset, N. Semaphorin 3A-Vascular Endothelial Growth Factor-165 Balance Mediates Migration and Apoptosis of Neural Progenitor Cells by the Recruitment of Shared Receptor. J. Neurosci. 2001, 21, 3332–3341. [Google Scholar] [CrossRef] [PubMed]
- Castro-Rivera, E.; Ran, S.; Thorpe, P.; Minna, J.D. Semaphorin 3B (SEMA3B) induces apoptosis in lung and breast cancer, whereas VEGF165 antagonizes this effect. Proc. Natl. Acad. Sci. USA 2004, 101, 11432–11437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Ding, M.; Qian, N.; Song, B.; Yu, J.; Tang, J.; Wang, J. Decreased expression of semaphorin 3D is associated with genesis and development in colorectal cancer. World J. Surg. Oncol. 2017, 15, 67–1128. [Google Scholar] [CrossRef]
- Kigel, B.; Varshavsky, A.; Kessler, O.; Neufeld, G. Successful inhibition of tumor development by specific class-3 semaphorins is associated with expression of appropriate semaphorin receptors by tumor cells. PLoS ONE 2008, 3, e3287. [Google Scholar] [CrossRef] [PubMed]
- Foley, K.; Rucki, A.A.; Xiao, Q.; Zhou, D.; Leubner, A.; Mo, G.; Kleponis, J.; Wu, A.A.; Sharma, R.; Jiang, Q.; et al. Semaphorin 3D autocrine signaling mediates the metastatic role of annexin A2 in pancreatic cancer. Sci. Signal. 2015, 8, ra77. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, M.; Avraham, I.; Dor, Y.; Bachar-Lustig, E.; Itin, A.; Yung, S.; Chimenti, S.; Landsman, L.; Abramovitch, R.; Keshet, E. VEGF-Induced Adult Neovascularization: Recruitment, Retention, and Role of Accessory Cells. Cell 2006, 124, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Zacchigna, S.; Pattarini, L.; Zentilin, L.; Moimas, S.; Carrer, A.; Sinigaglia, M.; Arsic, N.; Tafuro, S.; Sinagra, G.; Giacca, M. Bone marrow cells recruited through the neuropilin-1 receptor promote arterial formation at the sites of adult neoangiogenesis in mice. J. Clin. Investig. 2008, 118, 2062–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrer, A.; Moimas, S.; Zacchigna, S.; Pattarini, L.; Zentilin, L.; Ruozi, G.; Mano, M.; Sinigaglia, M.; Maione, F.; Serini, G.; et al. Neuropilin-1 Identifies a Subset of Bone Marrow Gr1- Monocytes That Can Induce Tumor Vessel Normalization and Inhibit Tumor Growth. Cancer Res. 2012, 15, 6371–6381. [Google Scholar] [CrossRef] [PubMed]
- Casazza, A.; Laoui, D.; Wenes, M.; Rizzolio, S.; Bassani, N.; Mambretti, M.; Deschoemaeker, S.; Van Ginderachter, J.A.; Tamagnone, L.; Mazzone, M. Impeding Macrophage Entry into Hypoxic Tumor Areas by Sema3A/Nrp1 Signaling Blockade Inhibits Angiogenesis and Restores Antitumor Immunity. Cancer Cell 2013, 24, 695–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolny, C.; Capparuccia, L.; Casazza, A.; Mazzone, M.; Vallario, A.; Cignetti, A.; Medico, E.; Carmeliet, P.; Comoglio, P.M.; Tamagnone, L. The tumor suppressor semaphorin 3B triggers a prometastatic program mediated by interleukin 8 and the tumor microenvironment. J. Exp. Med. 2008, 205, 1155–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Mills, L.; Mian, B.; Tellez, C.; Mccarty, M.; Yang, X.D.; Gudas, J.M.; Bar-Eli, M. Fully humanized neutralizing antibodies to interleukin-8 (ABX-IL8) inhibit angiogenesis, tumor growth, and metastasis of human melanoma. Am. J. Pathol. 2002, 161, 125–134. [Google Scholar] [CrossRef]
- Podgrabinska, S.; Skobe, M. Role of lymphatic vasculature in regional and distant metastases. Microvasc. Res. 2014, 95, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, S.; Detmar, M. Mechanisms of lymphatic metastasis. J. Clin. Investig. 2014, 124, 922–928. [Google Scholar] [CrossRef] [Green Version]
- Doci, C.L.; Mikelis, C.M.; Lionakis, M.S.; Molinolo, A.A.; Gutkind, J.S. Genetic identification of SEMA3F as an anti-lymphangiogenic metastasis suppressor gene in head and neck squamous carcinoma. Cancer Res. 2015, 75, 2937–2948. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Gao, Z.; Sun, M.; Li, H.; Fan, H.; Chen, D.; Zheng, J. Prognostic significance of VEGF-C, semaphorin 3F, and neuropilin-2 expression in oral squamous cell carcinomas and their relationship with lymphangiogenesis. J. Surg. Oncol. 2014, 111, 382–388. [Google Scholar] [CrossRef]
- Mucka, P.; Levonyak, N.; Geretti, E.; Zwaans, B.M.; Li, X.; Adini, I.; Klagsbrun, M.; Adam, R.M.; Bielenberg, D.R. Inflammation and Lymphedema Are Exacerbated and Prolonged by Neuropilin 2 Deficiency. Am. J. Pathol. 2016, 186, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; James, J.M.; Suto, F.; Mukouyama, Y.S. Class 3 semaphorins negatively regulate dermal lymphatic network formation. Biol. Open 2015, 4, 1194–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachelder, R.E.; Lipscomb, E.A.; Lin, X.; Wendt, M.A.; Chadborn, N.H.; Eickholt, B.J.; Mercurio, A.M. Competing Autocrine Pathways Involving Alternative Neuropilin-1 Ligands Regulate Chemotaxis of Carcinoma Cells. Cancer Res. 2003, 63, 5230–5233. [Google Scholar] [PubMed]
- Herman, J.G.; Meadows, G.G. Increased class 3 semaphorin expression modulates the invasive and adhesive properties of prostate cancer cells. Int. J. Oncol. 2007, 30, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Thorat, D.; Soundararajan, G.; Pradhan, S.J.; Chakraborty, G.; Lohite, K.; Karnik, S.; Kundu, G.C. Semaphorin 3A upregulates FOXO 3a-dependent MelCAM expression leading to attenuation of breast tumor growth and angiogenesis. Oncogene 2015, 34, 1584–1595. [Google Scholar] [CrossRef] [PubMed]
- Nehil, M.; Paquette, J.; Tokuyasu, T.; McCormick, F. High mobility group box 1 promotes tumor cell migration through epigenetic silencing of semaphorin 3A. Oncogene 2013, 33, 5151–5162. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, Q.; Yin, D.; Shi, S.; Yu, L.; Zhou, S.; Chen, E.; Zhou, Z.; Shi, Y.; Fan, J.; et al. Novel role of semaphorin 3A in the growth and progression of hepatocellular carcinoma. Oncol. Rep. 2017, 37, 3313–3320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagci, T.; Wu, J.K.; Pfannl, R.; Ilag, L.L.; Jay, D.G. Autocrine semaphorin 3A signaling promotes glioblastoma dispersal. Oncogene 2009, 28, 3537–3550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.W.; Giese, N.A.; Swiercz, J.M.; Ceyhan, G.O.; Esposito, I.; Hinz, U.; Buchler, P.; Giese, T.; Buchler, M.W.; Offermanns, S.; et al. Association of axon guidance factor semaphorin 3A with poor outcome in pancreatic cancer. Int. J. Cancer 2007, 121, 2421–2433. [Google Scholar] [CrossRef] [PubMed]
- Tse, C.; Xiang, R.H.; Bracht, T.; Naylor, S.L. Human Semaphorin 3B (SEMA3B) Located at Chromosome 3p21.3 Suppresses Tumor Formation in an Adenocarcinoma Cell Line. Cancer Res. 2002, 62, 542–546. [Google Scholar] [PubMed]
- Loginov, V.I.; Dmitriev, A.A.; Senchenko, V.N.; Pronina, I.V.; Khodyrev, D.S.; Kudryavtseva, A.V.; Krasnov, G.S.; Gerashchenko, G.V.; Chashchina, L.I.; Kazubskaya, T.P.; et al. Tumor Suppressor Function of the SEMA3B Gene in Human Lung and Renal Cancers. PLoS ONE 2015, 10, e0123369. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Ivanova, V.S.; Kavandi, L.; Rodriguez, G.C.; Maxwell, G.; Syed, V. Progesterone and 1,25-Dihydroxyvitamin D3 Inhibit Endometrial Cancer Cell Growth by Upregulating Semaphorin 3B and Semaphorin 3F. Mol. Cancer Res. 2011, 9, 1479–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Ling, T.; Wu, H.; Zhang, J. Screening of candidate tumor-suppressor genes in 3p21.3 and investigation of the methylation of gene promoters in oral squamous cell carcinoma. Oncol. Rep. 2013, 29, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Shahi, P.; Wang, C.Y.; Chou, J.; Hagerling, C.; Gonzalez, V.H.; Ruderisch, A.; Yu, Y.; Lai, M.D.; Werb, Z. GATA3 targets semaphorin 3B in mammary epithelial cells to suppress breast cancer progression and metastasis. Oncogene 2017, 36, 5567–5575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, R.; Davalos, A.R.; Hensel, C.H.; Zhou, X.J.; Tse, C.; Naylor, S.L. Semaphorin 3F Gene from Human 3p21.3 Suppresses Tumor Formation in Nude Mice. Cancer Res. 2002, 62, 2637–2643. [Google Scholar] [PubMed]
- Liu, Y.; Li, R.; Yin, K.; Ren, G.; Zhang, Y. The crucial role of SEMA3F in suppressing the progression of oral squamous cell carcinoma. Cell. Mol. Biol. Lett. 2017, 22, 32. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.H.; Fu, W.J.; Cui, Y.H.; Guo, Q.N.; Zhou, Y. Downregulation of Semaphorin-3F is associated with poor prognostic significance in osteosarcoma patients. Am. J. Cancer Res. 2016, 6, 2252–2262. [Google Scholar] [PubMed]
- Gao, X.; Tang, C.; Shi, W.; Feng, S.; Qin, W.; Jiang, T.; Sun, Y. Semaphorin-3F functions as a tumor suppressor in colorectal cancer due to regulation by DNA methylation. Int. J. Clin. Exp. Pathol. 2015, 8, 12766–12774. [Google Scholar] [PubMed]
- Bollard, J.; Massoma, P.; Vercherat, C.; Blanc, M.; Lepinasse, F.; Gadot, N.; Couderc, C.; Poncet, G.; Walter, T.; Joly, M.O.; et al. The axon guidance molecule Semaphorin 3F is a negative regulator of tumor progression and proliferation in ileal neuroendocrine tumors. Oncotarget 2015, 6, 36731–36745. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, Q.; Yang, J.; Duan, G.; Ou, J.; Zhang, R.; Pan, F.; Peng, Q.; Tan, H.; Ping, Y.F.; et al. Axon Guiding Chemorepulsant Semaphorin-3F Inhibits Tumor Growth and Metastasis of Colorectal Carcinoma. Clin. Cancer Res. 2011, 17, 2702–2711. [Google Scholar] [CrossRef] [PubMed]
- Nasarre, P.; Kusy, S.; Constantin, B.; Castellani, V.; Drabkin, H.A.; Bagnard, D.; Roche, J. Semaphorin SEMA3F Has a Repulsing Activity on Breast Cancer Cells and Inhibits E-Cadherin-Mediated Cell Adhesion. Neoplasia 2005, 7, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Potiron, V.A.; Sharma, G.; Nasarre, P.; Clarhaut, J.A.; Augustin, H.G.; Gemmill, R.M.; Roche, J.; Drabkin, H.A. Semaphorin SEMA3F Affects Multiple Signaling Pathways in Lung Cancer Cells. Cancer Res. 2007, 67, 8708–8715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusy, S.; Nasarre, P.; Chan, D.; Potiron, V.; Meyronet, D.; Gemmill, R.M.; Constantin, B.; Drabkin, H.A.; Roche, J. Selective suppression of in vivo tumorigenicity by semaphorin SEMA3F in lung cancer cells. Neoplasia 2005, 7, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Zhou, Z.; Yang, J.; Shi, Y.; Xu, S.; Wang, B.; Ping, Y.; Chen, L.; Cui, Y.; Zhang, X.; et al. Semaphorin-3F suppresses the stemness of colorectal cancer cells by inactivating Rac1. Cancer Lett. 2014, 358, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Endo, R.; Gotoh, M.; Hirohashi, S. Identification of semaphorin E as a non-MDR drug resistance gene of human cancers. Proc. Natl. Acad. Sci. USA 1997, 94, 14713–14718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaitkiene, P.; Skiriute, D.; Steponaitis, G.; Skauminas, K.; Tamasauskas, A.; Kazlauskas, A. High level of Sema3C is associated with glioma malignancy. Diagn. Pathol. 2015, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Man, J.; Shoemake, J.; Zhou, W.; Fang, X.; Wu, Q.; Rizzo, A.; Prayson, R.; Bao, S.; Rich, J.N.; Yu, J.S. Sema3C promotes the survival and tumorigenicity of glioma stem cells through Rac1 activation. Cell Rep. 2014, 9, 1812–1826. [Google Scholar] [CrossRef] [PubMed]
- Miyato, H.; Tsuno, N.H.; Kitayama, J. Semaphorin 3C is involved in the progression of gastric cancer. Cancer Sci. 2012, 103, 1961–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, V.; Nariculam, J.; Munson, P.; Freeman, A.; Klocker, H.; Masters, J.; Williamson, M. A role for class 3 semaphorins in prostate cancer. Prostate 2010, 71, 649–658. [Google Scholar] [CrossRef]
- Xu, X.; Zhao, Z.; Guo, S.; Li, J.; Liu, S.; You, Y.; Ni, B.; Wang, H.; Bie, P. Increased semaphorin 3c expression promotes tumor growth and metastasis in pancreatic ductal adenocarcinoma by activating the ERK1/2 signaling pathway. Cancer Lett. 2017, 397, 12–22. [Google Scholar] [CrossRef]
- Tam, K.J.; Hui, D.H.F.; Lee, W.W.; Dong, M.; Tombe, T.; Jiao, I.Z.F.; Khosravi, S.; Takeuchi, A.; Peacock, J.W.; Ivanova, L.; et al. Semaphorin 3 C drives epithelial-to-mesenchymal transition, invasiveness, and stem-like characteristics in prostate cells. Sci. Rep. 2017, 7, 11501–11914. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, X.; Ye, Z.; Chen, Y.; Lv, L.; Zhang, X.; Hu, H. Silencing of semaphorin 3C suppresses cell proliferation and migration in MCF-7 breast cancer cells. Oncol. Lett. 2017, 14, 5913–5917. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Yu, J.S. Semaphorin 3C and Its Receptors in Cancer and Cancer Stem-Like Cells. Biomedicines 2018, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Esselens, C.; Malapeira, J.; Colome, N.; Casal, C.; Rodriguez-Manzaneque, J.C.; Canals, F.; Arribas, J. The cleavage of semaphorin 3C induced by ADAMTS1 promotes cell migration. J. Biol. Chem. 2009, 285, 2463–2473. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.E.; Fu, J.; Lopez de Cicco, R.; Klein-Szanto, A.J. Proprotein convertases: “Master switches” in the regulation of tumor growth and progression. Mol. Carcinog. 2005, 44, 151–161. [Google Scholar] [CrossRef]
- Zhou, X.; Ma, L.; Li, J.; Gu, J.; Shi, Q.; Yu, R. Effects of SEMA3G on migration and invasion of glioma cells. Oncol. Rep. 2012, 28, 269–275. [Google Scholar] [PubMed] [Green Version]
- Roodink, I.; Kats, G.; van Kempen, L.; Grunberg, M.; Maass, C.; Verrijp, K.; Raats, J.; Leenders, W. Semaphorin 3E Expression Correlates Inversely with Plexin D1 During Tumor Progression. Am. J. Pathol. 2008, 173, 1873–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, C.R.; Klingelhofer, J.; Tarabykina, S.; Hulgaard, E.F.; Kramerov, D.; Lukanidin, E. Transcription of a novel mouse semaphorin gene, M-semaH, correlates with the metastatic ability of mouse tumor cell lines. Cancer Res. 1998, 58, 1238–1244. [Google Scholar] [PubMed]
- Schmidt, A.M.; Moore, K.J. The Semaphorin 3E/PlexinD1 Axis Regulates Macrophage Inflammation in Obesity. Cell Metab. 2013, 18, 461–462. [Google Scholar] [CrossRef] [Green Version]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben Neriah, Y. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef]
- Sierra, J.R.; Corso, S.; Caione, L.; Cepero, V.; Conrotto, P.; Cignetti, A.; Piacibello, W.; Kumanogoh, A.; Kikutani, H.; Comoglio, P.M.; et al. Tumor angiogenesis and progression are enhanced by Sema4D produced by tumor-associated macrophages. J. Exp. Med. 2008, 205, 1673–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Sica, A. Macrophages, innate immunity and cancer: Balance, tolerance, and diversity. Curr. Opin. Immunol. 2010, 22, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Catalano, A.; Caprari, P.; Moretti, S.; Faronato, M.; Tamagnone, L.; Procopio, A. Semaphorin-3A is expressed by tumor cells and alters T-cell signal transduction and function. Blood 2006, 107, 3321–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Suzuki, K.; Okuno, T.; Ogata, T.; Takegahara, N.; Takamatsu, H.; Mizui, M.; Taniguchi, M.; Chedotal, A.; Suto, F.; et al. Plexin-A4 negatively regulates T lymphocyte responses. Int. Immunol. 2008, 20, 413–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallerius, M.; Wallmann, T.; Bartish, M.; Ostling, J.; Mezheyeuski, A.; Tobin, N.P.; Nygren, E.; Pangigadde, P.; Pellegrini, P.; Squadrito, M.L.; et al. Guidance Molecule SEMA3A Restricts Tumor Growth by Differentially Regulating the Proliferation of Tumor-Associated Macrophages. Cancer Res. 2016, 76, 3166–3178. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Da-Cruz, D.A.; Brignier, A.C.; Asnafi, V.; Baleydier, F.; Messias, C.V.; Lepelletier, Y.; Bedjaoui, N.; Renand, A.; Smaniotto, S.; Canioni, D.; et al. Semaphorin 3F and neuropilin-2 control the migration of human T-cell precursors. PLoS ONE 2014, 9, e103405. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, H.; Koussih, L.; Shan, L.; Gounni, A.S. The Regulatory Role of Semaphorin 3E in Allergic Asthma. Int. J. Biochem. Cell Biol. 2018, 106, 68–73. [Google Scholar] [CrossRef]
- Curreli, S.; Wong, B.S.; Latinovic, O.; Konstantopoulos, K.; Stamatos, N.M. Class 3 semaphorins induce F-actin reorganization in human dendritic cells: Role in cell migration. J. Leukoc. Biol. 2016, 100, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Sekido, Y.; Bader, S.; Latif, F.; Chen, J.Y.; Duh, F.M.; Wei, M.H.; Albanesi, J.P.; Lee, C.C.; Lerman, M.I.; Minna, J.D. Human semaphorins A(V) and IV reside in the 3p21.3 small cell lung cancer deletion region and demonstrate distinct expression patterns. Proc. Natl. Acad. Sci. USA 1996, 93, 4120–4125. [Google Scholar] [CrossRef]
- Lerman, M.I.; Minna, J.D. The 630-kb lung cancer homozygous deletion region on human chromosome 3p21.3: Identification and evaluation of the resident candidate tumor suppressor genes. Cancer Res. 2000, 60, 6116–6133. [Google Scholar]
- Kuroki, T.; Trapasso, F.; Yendamuri, S.; Matsuyama, A.; Alder, H.; Williams, N.N.; Kaiser, L.R.; Croce, C.M. Allelic Loss on Chromosome 3p21.3 and Promoter Hypermethylation of Semaphorin 3B in Non-Small Cell Lung Cancer. Cancer Res. 2003, 63, 3352–3355. [Google Scholar]
- Tischoff, I.; Markwarth, A.; Witzigmann, H.; Uhlmann, D.; Hauss, J.; Mirmohammadsadegh, A.; Wittekind, C.; Hengge, U.R.; Tannapfel, A. Allele loss and epigenetic inactivation of 3p21.3 in malignant liver tumors. Int. J. Cancer 2005, 115, 684–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, P.N.; McArdle, L.; Cornell, J.; Cohn, S.L.; Stallings, R.L. High-resolution analysis of 3p deletion in neuroblastoma and differential methylation of the SEMA3B tumor suppressor gene. Cancer Genet. Cytogenet. 2007, 174, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.H.; Du, W.; Long, J.; Song, L.J. Mechanism of SEMA3B gene silencing and clinical significance in glioma. Genet. Mol. Res. 2016, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, E.; Tang, M.; Baez, S.; Diaz, A.; Pruyas, M.; Wistuba, I.I.; Corvalan, A. Frequent epigenetic inactivation of chromosome 3p candidate tumor suppressor genes in gallbladder carcinoma. Cancer Lett. 2007, 250, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Yim, S.H.; Hu, H.J.; Jung, C.K.; Lee, S.H.; Kim, D.H.; Chung, Y.J. Copy number alterations and expression profiles of candidate genes in a pulmonary inflammatory myofibroblastic tumor. Lung Cancer 2010, 70, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Marsit, C.J.; Wiencke, J.K.; Liu, M.; Kelsey, K.T. The race associated allele of semaphorin 3B (SEMA3B) T415I and its role in lung cancer in African-Americans and Latino-Americans. Carcinogenesis 2005, 26, 1446–1449. [Google Scholar] [CrossRef] [PubMed]
- Michikawa, Y.; Suga, T.; Ishikawa, A.; Hayashi, H.; Oka, A.; Inoko, H.; Iwakawa, M.; Imai, T. Genome wide screen identifies microsatellite markers associated with acute adverse effects following radiotherapy in cancer patients. BMC Med. Genet. 2010, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shin, Y.J.; Lee, K.; Cho, H.J.; Sa, J.K.; Lee, S.Y.; Kim, S.H.; Lee, J.; Yoon, Y.; Nam, D.H. Anti-SEMA3A Antibody: A Novel Therapeutic Agent to Suppress GBM Tumor Growth. Cancer Res. Treat. 2018, 50, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Sorber, R.; Teper, Y.; bisoye-Ogunniyan, A.; Waterfall, J.J.; Davis, S.; Killian, J.K.; Pineda, M.; Ray, S.; McCord, M.R.; Pflicke, H.; et al. Whole Genome Sequencing of Newly Established Pancreatic Cancer Lines Identifies Novel Somatic Mutation (c.2587G>A) in Axon Guidance Receptor Plexin A1 as Enhancer of Proliferation and Invasion. PLoS ONE 2016, 11, e0149833. [Google Scholar] [CrossRef] [PubMed]
- Villacis, R.A.; Miranda, P.M.; Gomy, I.; Santos, E.M.; Carraro, D.M.; Achatz, M.I.; Rossi, B.M.; Rogatto, S.R. Contribution of rare germline copy number variations and common susceptibility loci in Lynch syndrome patients negative for mutations in the mismatch repair genes. Int. J. Cancer 2016, 138, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, Y.; Sato, A.; Tsujimura, T.; Morinaga, T.; Fukuoka, K.; Yamada, S.; Murakami, A.; Kondo, N.; Matsumoto, S.; Okumura, Y.; et al. Frequent deletion of 3p21.1 region carrying semaphorin 3G and aberrant expression of the genes participating in semaphorin signaling in the epithelioid type of malignant mesothelioma cells. Int. J. Oncol. 2011, 39, 1365–1374. [Google Scholar] [PubMed]
- Meyer, L.A.; Fritz, J.; Pierdant-Mancera, M.; Bagnard, D. Current drug design to target the Semaphorin/Neuropilin/Plexin complexes. Cell Adhes. Migr. 2016, 10, 700–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledano, S.; Nir-Zvi, I.; Engelman, R.; Kessler, O.; Neufeld, G. Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression. Int. J. Mol. Sci. 2019, 20, 556. https://doi.org/10.3390/ijms20030556
Toledano S, Nir-Zvi I, Engelman R, Kessler O, Neufeld G. Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression. International Journal of Molecular Sciences. 2019; 20(3):556. https://doi.org/10.3390/ijms20030556
Chicago/Turabian StyleToledano, Shira, Inbal Nir-Zvi, Rotem Engelman, Ofra Kessler, and Gera Neufeld. 2019. "Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression" International Journal of Molecular Sciences 20, no. 3: 556. https://doi.org/10.3390/ijms20030556