
Key Points
-
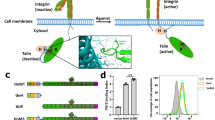
Integrins are transmembrane receptors that support adhesion to the extracellular matrix and transmit chemical and mechanical signals into and out of cells. Cells regulate integrin-mediated adhesion by controlling integrin affinity for extracellular ligands (integrin activation), by clustering integrins into adhesion complexes (avidity modulation) and by regulating links between integrin cytoplasmic domains and the actin cytoskeleton.
-
The large dimeric actin-binding protein talin is a key regulator of integrin activation, and is composed of an amino-terminal head and a carboxy-terminal flexible rod. The structures of all 18 domains in talin have now been determined, and a model of full-length talin has been generated.
-
Biochemical and structural data reveal how direct interactions between the short cytoplasmic tails of β-integrin subunits and the talin head (a FERM domain) disrupt inhibitory interactions between the integrin α-subunit and β-subunit, inducing conformational changes in the integrin extracellular domains that result in their increased affinity for ligand.
-
The talin rod consists of 13 either 4- or 5-helix bundles, terminating in a single helix that supports talin dimerization. It contains two actin-binding sites and multiple binding sites for the actin-binding protein vinculin, which stabilizes adhesion complexes. Recent data show that the dynamic between talin–RIAM and talin–vinculin interactions is important for integrin activation and force-induced maturation of adhesions.
-
Regulation of talin–integrin interactions represents an important control point along integrin activation pathways. Various regulatory mechanisms have been identified, including talin autoinhibition, competition between talin and other integrin tail-binding or talin-binding proteins and post-translational modification of β-integrin subunits.
-
Although much less well-understood than talins, kindlin family proteins are also known to be important integrin regulators. Kindlin loss, due to knockout, knockdown or disease-causing mutations, result in defective integrin activation.
-
The molecular basis of kindlin function has yet to be determined, but kindlins are structurally related to the talin head, directly bind β-integrin tails and membranes, localize to focal adhesions and synergize with talin during integrin activation.
Abstract
Integrin receptors provide a dynamic, tightly-regulated link between the extracellular matrix (or cellular counter-receptors) and intracellular cytoskeletal and signalling networks, enabling cells to sense and respond to their chemical and physical environment. Talins and kindlins, two families of FERM–domain proteins, bind the cytoplasmic tail of integrins, recruit cytoskeletal and signalling proteins involved in mechanotransduction and synergize to activate integrin binding to extracellular ligands. New data reveal the domain structure of full-length talin, provide insights into talin-mediated integrin activation and show that RIAM recruits talin to the plasma membrane, whereas vinculin stabilizes talin in cell–matrix junctions. How kindlins act is less well-defined, but disease-causing mutations show that kindlins are also essential for integrin activation, adhesion, cell spreading and signalling.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


References
Burridge, K. & Connell, L. A. New protein of adhesion plaques and ruffling membranes. J. Cell Biol. 97, 359–367 (1983).
Critchley, D. R. Biochemical and structural properties of the integrin-associated cytoskeletal protein talin. Annu. Rev. Biophys. 38, 235–254 (2009).
Calderwood, D. A. et al. The talin head domain binds to integrin-β subunit cytoplasmic tails and regulates integrin activation. J. Biol. Chem. 274, 28071–28704 (1999).
Tadokoro, S. et al. Talin binding to integrin-β tails: a final common step in integrin activation. Science 302, 103–106 (2003).
Garcia-Alvarez, B. et al. Structural determinants of integrin recognition by talin. Mol. Cell 11, 49–58 (2003).
Wegener, K. L. et al. Structural basis of integrin activation by talin. Cell 128, 171–182 (2007).
Calderwood, D. A. Integrin activation. J. Cell Sci. 117, 657–666 (2004).
Brown, N. H. et al. Talin is essential for integrin function in Drosophila. Dev. Cell 3, 569–579 (2002).
Cram, E. J., Clark, S. G. & Schwarzbauer, J. E. Talin loss-of-function uncovers roles in cell contractility and migration in C. elegans. J. Cell Sci. 116, 3871–3878 (2003).
Zhang, X. et al. Talin depletion reveals independence of initial cell spreading from integrin activation and traction. Nature Cell Biol. 10, 1062–1068 (2008).
Kopp, P. M. et al. Studies on the morphology and spreading of human endothelial cells define key inter- and intramolecular interactions for talin1. Eur. J. Cell Biol. 89, 661–673 (2010).
Monkley, S. J. et al. Endothelial cell talin1 is essential for embryonic angiogenesis. Dev. Biol. 349, 494–502 (2011).
Moser, M., Nieswandt, B., Ussar, S., Pozgajova, M. & Fassler, R. Kindlin-3 is essential for integrin activation and platelet aggregation. Nature Med. 14, 325–330 (2008). First demonstration that loss of kindlin 3 results in severe defects in platelet integrin activation, revealing the importance of kindlins in integrin activation.
Rogalski, T. M., Mullen, G. P., Gilbert, M. M., Williams, B. D. & Moerman, D. G. The unc-112 gene in Caenorhabditis elegans encodes a novel component of cell–matrix adhesion structures required for integrin localization in the muscle cell membrane. J. Cell Biol. 150, 253–264 (2000).
Siegel, D. H. et al. Loss of kindlin-1, a human homolog of the Caenorhabditis elegans actin–extracellular-matrix linker protein UNC-112, causes Kindler syndrome. Am. J. Hum. Genet. 73, 174–187 (2003).
Jobard, F. et al. Identification of mutations in a new gene encoding a FERM family protein with a pleckstrin homology domain in Kindler syndrome. Hum. Mol. Genet. 12, 925–935 (2003).
Karakose, E., Schiller, H. B. & Fassler, R. The kindlins at a glance. J. Cell Sci. 123, 2353–2356 (2010).
Kloeker, S. et al. The Kindler syndrome protein is regulated by transforming growth factor-β and involved in integrin-mediated adhesion. J. Biol. Chem. 279, 6824–6833 (2004).
Plow, E. F., Qin, J. & Byzova, T. Kindling the flame of integrin activation and function with kindlins. Curr. Opin. Hematol. 16, 323–328 (2009).
Ye, F., Kim, C. & Ginsberg, M. H. Molecular mechanism of inside–out integrin regulation. J. Thromb. Haemost. 9 (Suppl. 1), 20–25 (2011).
Anthis, N. J. & Campbell, I. D. The tail of integrin activation. Trends Biochem. Sci. 36, 191–198 (2011).
Shattil, S. J., Kim, C. & Ginsberg, M. H. The final steps of integrin activation: the end game. Nature Rev. Mol. Cell Biol. 11, 288–300 (2010).
Peng, X., Nelson, E. S., Maiers, J. L. & DeMali, K. A. New insights into vinculin function and regulation. Int. Rev. Cell. Mol. Biol. 287, 191–231 (2011).
Ziegler, W. H., Liddington, R. C. & Critchley, D. R. The structure and regulation of vinculin. Trends Cell Biol. 16, 453–460 (2006).
Winkler, J., Lunsdorf, H. & Jockusch, B. M. Energy-filtered electron microscopy reveals that talin is a highly flexible protein composed of a series of globular domains. Eur. J. Biochem. 243, 430–436 (1997).
Elliott, P. R. et al. The structure of the talin head reveals a novel extended conformation of the FERM domain. Structure 18, 1289–1299 (2010).
Gingras, A. R. et al. The structure of the C-terminal actin-binding domain of talin. EMBO J. 27, 458–469 (2008).
Gingras, A. R. et al. Central region of talin has a unique fold that binds vinculin and actin. J. Biol. Chem. 285, 29577–29587 (2010).
Gingras, A. R. et al. Structural and dynamic characterization of a vinculin binding site in the talin rod. Biochemistry 45, 1805–1817 (2006).
Gingras, A. R. et al. Structural determinants of integrin binding to the talin rod. J. Biol. Chem. 284, 8866–8876 (2009).
Gingras, A. R. et al. Mapping and consensus sequence identification for multiple vinculin binding sites within the talin rod. J. Biol. Chem. 280, 37217–37224 (2005).
Goult, B. T. et al. Structure of a double ubiquitin-like domain in the talin head: a role in integrin activation. EMBO J. 29, 1069–1080 (2010).
Goult, B. T. et al. The domain structure of talin: residues 1815–1973 form a five-helix bundle containing a cryptic vinculin-binding site. FEBS Lett. 584, 2237–2241 (2010).
Papagrigoriou, E. et al. Activation of a vinculin-binding site in the talin rod involves rearrangement of a five-helix bundle. EMBO J. 23, 2942–2951 (2004).
Goult, B. T. et al. RIAM and vinculin binding to talin are mutually exclusive and regulate adhesion assembly and turnover. J. Biol. Chem. 288, 8238–8249 (2013). Presents a structural model for full-length talin and provides a structural basis for a switch from talin–RIAM to talin–vinculin complexes during adhesion assembly and maturation.
Anthis, N. J., Wegener, K. L., Critchley, D. R. & Campbell, I. D. Structural diversity in integrin/talin interactions. Structure 18, 1654–1666 (2010).
Anthis, N. J. et al. The structure of an integrin/talin complex reveals the basis of inside–out signal transduction. EMBO J. 28, 3623–3632 (2009). The first structure of a full-length β-integrin tail bound to the talin F2–F3 FERM domains. This, combined with biophysical studies and integrin activation assays, reveals that talin F3 binding to the membrane-proximal helix of the β-integrin tail disrupts an inhibitory α-integrin tail–β-integrin tail interaction, and identifies an important positively charged membrane-binding surface on the talin F2 domain.
Wegener, K. L. et al. Structural basis for the interaction between the cytoplasmic domain of the hyaluronate receptor layilin and the talin F3 subdomain. J. Mol. Biol. 382, 112–126 (2008).
de Pereda, J. M. et al. Structural basis for phosphatidylinositol phosphate kinase type Iγ binding to talin at focal adhesions. J. Biol. Chem. 280, 8381–8386 (2005).
Lawson, C. et al. FAK promotes recruitment of talin to nascent adhesions to control cell motility. J. Cell Biol. 196, 223–232 (2012).
Wang, S. et al. Tiam1 interaction with the PAR complex promotes talin-mediated Rac1 activation during polarized cell migration. J. Cell Biol. 199, 331–345 (2012).
Bate, N. et al. Talin contains a C-terminal calpain2 cleavage site important in focal adhesion dynamics. PLoS ONE 7, e34461 (2012).
Rodius, S. et al. The talin rod IBS2 α-helix interacts with the β3 integrin cytoplasmic tail membrane-proximal helix by establishing charge complementary salt bridges. J. Biol. Chem. 283, 24212–24223 (2008).
Han, J. et al. Reconstructing and deconstructing agonist-induced activation of integrin αIIbβ3. Curr. Biol. 16, 1796–1806 (2006).
Lee, H. S., Lim, C. J., Puzon-McLaughlin, W., Shattil, S. J. & Ginsberg, M. H. RIAM activates integrins by linking talin to Ras GTPase membrane-targeting sequences. J. Biol. Chem. 284, 5119–5127 (2009). Provides the first insights into how the RAP1A effector RIAM binds to talin and recruits it to the membrane to activate integrins.
Watanabe, N. et al. Mechanisms and consequences of agonist-induced talin recruitment to platelet integrin αIIbβ3. J. Cell Biol. 181, 1211–1222 (2008).
Li, G. et al. Full activity of the deleted in liver cancer 1 (DLC1) tumor suppressor depends on an LD-like motif that binds talin and focal adhesion kinase (FAK). Proc. Natl Acad. Sci. USA 108, 17129–17134 (2011).
Sun, N., Critchley, D. R., Paulin, D., Li, Z. & Robson, R. M. Identification of a repeated domain within mammalian α-synemin that interacts directly with talin. Exp. Cell Res. 314, 1839–1849 (2008).
Petrich, B. G. Talin-dependent integrin signalling in vivo. Thromb. Haemost. 101, 1020–1024 (2009).
Lau, T. L., Kim, C., Ginsberg, M. H. & Ulmer, T. S. The structure of the integrin αIIbβ3 transmembrane complex explains integrin transmembrane signalling. EMBO J. 28, 1351–1361 (2009). An NMR structure that provides key information about the complex formed by the membrane-spanning helices in the inactive integrin state.
Zhu, J. et al. The structure of a receptor with two associating transmembrane domains on the cell surface: integrin αIIbβ3. Mol. Cell 34, 234–249 (2009). A structure of the membrane-spanning region of an intact integrin, determined by disulfide crosslinking and molecular modelling.
Kim, C. et al. Basic amino-acid side chains regulate transmembrane integrin signalling. Nature 481, 209–213 (2011).
Kalli, A. C., Campbell, I. D. & Sansom, M. S. P. Multiscale simulations suggest a mechanism for integrin inside–out activation. Proc. Natl Acad. Sci. USA 108, 11890–11895 (2011).
Kim, C., Ye, F., Hu, X. & Ginsberg, M. H. Talin activates integrins by altering the topology of the β-transmembrane domain. J. Cell Biol. 197, 605–611 (2012). Demonstrates, together with reference 52, using environment-sensitive fluorophores, that talin binding to the β-integrin TMD alters the membrane embedding of this region.
Moore, D. T. et al. Affinity of talin-1 for the β3-integrin cytosolic domain is modulated by its phospholipid bilayer environment. Proc. Natl Acad. Sci. USA 109, 793–798 (2012).
Lefort, C. T. et al. Distinct roles for talin-1 and kindlin-3 in LFA-1 extension and affinity regulation. Blood 119, 4275–4282 (2012).
Nieswandt, B. et al. Loss of talin1 in platelets abrogates integrin activation, platelet aggregation, and thrombus formation in vitro and in vivo. J. Exp. Med. 204, 3113–3118 (2007).
Petrich, B. G. et al. Talin is required for integrin-mediated platelet function in hemostasis and thrombosis. J. Exp. Med. 204, 3103–3111 (2007).
Haling, J. R., Monkley, S. J., Critchley, D. R. & Petrich, B. G. Talin-dependent integrin activation is required for fibrin clot retraction by platelets. Blood 117, 1719–1722 (2011).
Kim, M., Carman, C. V. & Springer, T. A. Bidirectional transmembrane signaling by cytoplasmic domain separation in integrins. Science 301, 1720–1725 (2003).
Bouaouina, M., Harburger, D. S. & Calderwood, D. A. Talin and signaling through integrins. Methods Mol. Biol. 757, 325–347 (2012).
Goksoy, E. et al. Structural basis for the autoinhibition of talin in regulating integrin activation. Mol. Cell 31, 124–133 (2008). The first biochemical and structural data showing how binding of the talin head to the rod occludes the integrin-binding site in the head and how talin autoinhibition might be relieved by PtdIns(4,5)P 2.
Goult, B. et al. Structural studies on full-length talin1 reveal a compact auto-inhibited dimer: implications for talin activation. J. Struct. Biol. http://dx.doi.org/10.1016/j.jsb.2013.05.014 (2013).
Goult, B. T. et al. The structure of an interdomain complex that regulates talin activity. J. Biol. Chem. 284, 15097–15106 (2009).
Song, X. et al. A novel membrane-dependent on/off switch mechanism of talin FERM domain at sites of cell adhesion. Cell Res. 22, 1533–1545 (2012).
Saltel, F. et al. New PI(4,5)P2 and membrane proximal integrin-binding motifs in the talin head control β3-integrin clustering. J. Cell Biol. 187, 715–731 (2009).
Helsten, T. L. et al. Differences in regulation of Drosophila and vertebrate integrin affinity by talin. Mol. Biol. Cell 19, 3589–3598 (2008).
Bunch, T. A. Integrin αIIbβ3 activation in Chinese hamster ovary cells and platelets increases clustering rather than affinity. J. Biol. Chem. 285, 1841–1849 (2010).
Molony, L., Mccaslin, D., Abernethy, J., Paschal, B. & Burridge, K. Properties of talin from chicken gizzard smooth-muscle. J. Biol. Chem. 262, 7790–7795 (1987).
Banno, A. et al. Subcellular localization of talin is regulated by inter-domain interactions. J. Biol. Chem. 287, 13799–13812 (2012).
Bakolitsa, C. et al. Structural basis for vinculin activation at sites of cell adhesion. Nature 430, 583–586 (2004).
Humphries, J. D. et al. Vinculin controls focal adhesion formation by direct interactions with talin and actin. J. Cell Biol. 179, 1043–1057 (2007).
Carisey, A. et al. Vinculin regulates the recruitment and release of core focal adhesion proteins in a force-dependent manner. Curr. Biol. 23, 271–281 (2013).
Lee, H. S., Anekal, P., Lim, C. J., Liu, C. C. & Ginsberg, M. H. Two modes of integrin activation form a binary molecular switch in adhesion maturation. Mol. Biol. Cell 24, 1354–1362 (2013). Provides evidence that vinculin displaces RIAM from talin, and that this drives the transition from dynamic RIAM-positive nascent adhesions that support membrane protrusion to more stable vinculin-rich force-bearing focal adhesions.
Colo, G. P., Lafuente, E. M. & Teixido, J. The MRL proteins: adapting cell adhesion, migration and growth. Eur. J. Cell Biol. 91, 861–868 (2012).
Wynne, J. P. et al. Rap1-interacting adapter molecule (RIAM) associates with the plasma membrane via a proximity detector. J. Cell Biol. 199, 317–329 (2012).
Legate, K. R. et al. Integrin adhesion and force coupling are independently regulated by localized PtdIns(4,5)2 synthesis. EMBO J. 30, 4539–4553 (2011). Provides definitive evidence that PtdIns(4,5)P 2 generated by the talin-binding isoform of PtdInsP kinase Iγ in focal adhesions is essential for the formation of initial integrin-mediated cell attachment and for the subsequent linkage of integrins to cytoskeletal actin and the exertion of force on the matrix.
Thapa, N. et al. Phosphoinositide signaling regulates the exocyst complex and polarized integrin trafficking in directionally migrating cells. Dev. Cell 22, 116–130 (2012).
Li, X. et al. Ubiquitination of PIPKIγ90 by HECTD1 regulates focal adhesion dynamics and cell migration. J. Cell Sci. 126, 2617–2628 (2013).
Miller, N. L., Lawson, C., Chen, X. L., Lim, S. T. & Schlaepfer, D. D. Rgnef (p190RhoGEF) knockout inhibits RhoA activity, focal adhesion establishment, and cell motility downstream of integrins. PLoS ONE 7, e37830 (2012).
Rossier, O. et al. Integrins β1 and β3 exhibit distinct dynamic nanoscale organizations inside focal adhesions. Nature Cell Biol. 14, 1057–1067 (2012).
Wang, P., Ballestrem, C. & Streuli, C. H. The C-terminus of talin links integrins to cell cycle progression. J. Cell Biol. 195, 499–513 (2011).
Pouwels, J., Nevo, J., Pellinen, T., Ylanne, J. & Ivaska, J. Negative regulators of integrin activity. J. Cell Sci. 125, 3271–3280 (2012).
Anthis, N. J. et al. β-integrin tyrosine phosphorylation is a conserved mechanism for regulating talin-induced integrin activation. J. Biol. Chem. 284, 36700–36710 (2009).
Oxley, C. L. et al. An integrin phosphorylation switch: the effect of β3 integrin tail phosphorylation on Dok1 and talin binding. J. Biol. Chem. 283, 5420–5426 (2008).
Calderwood, D. A. et al. Integrin-β cytoplasmic domain interactions with phosphotyrosine-binding domains: a structural prototype for diversity in integrin signaling. Proc. Natl Acad. Sci. USA 100, 2272–2277 (2003).
Kiema, T. et al. The molecular basis of filamin binding to integrins and competition with talin. Mol. Cell 21, 337–347 (2006).
Ithychanda, S. S. et al. Migfilin, a molecular switch in regulation of integrin activation. J. Biol. Chem. 284, 4713–4722 (2009).
Lad, Y. et al. Structural basis of the migfilin–filamin interaction and competition with integrin-β tails. J. Biol. Chem. 283, 35154–35163 (2008).
Das, M., Ithychanda, S. S., Qin, J. & Plow, E. F. Migfilin and filamin as regulators of integrin activation in endothelial cells and neutrophils. PLoS ONE 6, e26355 (2011).
Moik, D. V., Janbandhu, V. C. & Fassler, R. Loss of migfilin expression has no overt consequences on murine development and homeostasis. J. Cell Sci. 124, 414–421 (2011).
Xiao, G. et al. Critical role of filamin-binding LIM protein 1 (FBLP-1)/migfilin in regulation of bone remodeling. J. Biol. Chem. 287, 21450–21460 (2012).
Takala, H. et al. β2 integrin phosphorylation on Thr758 acts as a molecular switch to regulate 14-3-3 and filamin binding. Blood 112, 1853–1862 (2008).
Millon-Fremillon, A. et al. Cell adaptive response to extracellular matrix density is controlled by ICAP-1-dependent β1-integrin affinity. J. Cell Biol. 180, 427–441 (2008).
Liu, W., Draheim, K. M., Zhang, R., Calderwood, D. A. & Boggon, T. J. Mechanism for KRIT1 release of ICAP1-mediated suppression of integrin activation. Mol. Cell 49, 719–729 (2013).
Roca-Cusachs, P. et al. Integrin-dependent force transmission to the extracellular matrix by α-actinin triggers adhesion maturation. Proc. Natl Acad. Sci. USA 110, e1361–e1370 (2013).
Tadokoro, S. et al. A potential role for α-actinin in inside–out αIIbβ3 signaling. Blood 117, 250–258 (2011).
Calderwood, D. A., Tai, V., Di Paolo, G., De Camilli, P. & Ginsberg, M. H. Competition for talin results in trans-dominant inhibition of integrin activation. J. Biol. Chem. 279, 28889–28895 (2004).
Bouaouina, M. & Calderwood, D. A. Kindlins. Curr. Biol. 21, R99–R101 (2011).
Ye, F. & Petrich, B. G. Kindlin: helper, co-activator, or booster of talin in integrin activation? Curr. Opin. Hematol. 18, 356–360 (2011).
Harburger, D. S., Bouaouina, M. & Calderwood, D. A. Kindlin-1 and -2 directly bind the C-terminal region of β-integrin cytoplasmic tails and exert integrin-specific activation effects. J. Biol. Chem. 284, 11485–11497 (2009).
Ma, Y. Q., Qin, J., Wu, C. & Plow, E. F. Kindlin-2 (Mig-2): a co-activator of β3 integrins. J. Cell Biol. 181, 439–446 (2008). Provides evidence for the cooperativity between the talin head and kindlin 2 in integrin activation.
Montanez, E. et al. Kindlin-2 controls bidirectional signaling of integrins. Genes Dev. 22, 1325–1330 (2008).
Malinin, N. L. et al. A point mutation in KINDLIN3 ablates activation of three integrin subfamilies in humans. Nature Med. 15, 313–318 (2009).
Qu, H. et al. Kindlin-2 regulates podocyte adhesion and fibronectin matrix deposition through interactions with phosphoinositides and integrins. J. Cell Sci. 124, 879–891 (2011).
Bledzka, K. et al. Spatial coordination of kindlin-2 with talin head domain in interaction with integrin-β cytoplasmic tails. J. Biol. Chem. 287, 24585–24594 (2012).
Ye, F. et al. Recreation of the terminal events in physiological integrin activation. J. Cell Biol. 188, 157–173 (2010). Establishes that purified talin components, in an in vitro nanodisc system, are sufficient to activate isolated membrane-embedded integrins, even in the absence of kindlin. Also provides support for a model whereby integrins adopt a more upright orientation when activated.
Goult, B. T. et al. The structure of the N-terminus of kindlin-1: a domain important for αIIbβ3 integrin activation. J. Mol. Biol. 394, 944–956 (2009).
Perera, H. D. et al. Membrane binding of the N-terminal ubiquitin-like domain of kindlin-2 is crucial for its regulation of integrin activation. Structure 19, 1664–1671 (2011).
Liu, J. et al. Structural basis of phosphoinositide binding to kindlin-2 protein pleckstrin homology domain in regulating integrin activation. J. Biol. Chem. 286, 43334–43342 (2011).
Liu, Y., Zhu, Y., Ye, S. & Zhang, R. Crystal structure of kindlin-2 PH domain reveals a conformational transition for its membrane anchoring and regulation of integrin activation. Protein Cell 3, 434–440 (2012).
Yates, L. A. et al. Structural and functional characterisation of the kindlin-1 pleckstrin homology domain. J. Biol. Chem. 287, 43246–43261 (2012).
Yates, L. A., Fuzery, A. K., Bonet, R., Campbell, I. D. & Gilbert, R. J. C. Biophysical analysis of kindlin-3 reveals an elongated conformation and maps integrin binding to the membrane-distal β-subunit, NPXY motif. J. Biol. Chem. 287, 37715–37731 (2012).
Moser, M., Legate, K. R., Zent, R. & Fassler, R. The tail of integrins, talin, and kindlins. Science 324, 895–899 (2009).
Kahner, B. N. et al. Kindlins, integrin activation and the regulation of talin recruitment to αIIbβ3. PLoS ONE 7, e34056 (2012).
Bouaouina, M. et al. A conserved lipid-binding loop in the kindlin FERM F1 domain is required for kindlin-mediated αIIbβ3 integrin coactivation. J. Biol. Chem. 287, 6979–6990 (2012).
Honda, S. et al. Integrin-linked kinase associated with integrin activation. Blood 113, 5304–5313 (2009).
Bandyopadhyay, A., Rothschild, G., Kim, S., Calderwood, D. A. & Raghavan, S. Functional differences between kindlin-1 and kindlin-2 in keratinocytes. J. Cell Sci. 125, 2172–2184 (2012).
Bialkowska, K. et al. The integrin co-activator Kindlin-3 is expressed and functional in a non-hematopoietic cell, the endothelial cell. J. Biol. Chem. 285, 18640–18649 (2010).
Ussar, S. et al. Loss of Kindlin-1 causes skin atrophy and lethal neonatal intestinal epithelial dysfunction. PLoS Genet. 4, e1000289 (2008).
Roca-Cusachs, P., Gauthier, N. C., Del Rio, A. & Sheetz, M. P. Clustering of α5β1 integrins determines adhesion strength whereas αvβ3 and talin enable mechanotransduction. Proc. Natl Acad. Sci. USA 106, 16245–16250 (2009).
del Rio, A. et al. Stretching single talin rod molecules activates vinculin binding. Science 323, 638–641 (2009). Use of magnetic tweezers, total internal reflection fluorescence and atomic force microscopy showed that physiologically relevant forces cause stretching of talin and result in exposure of cryptic VBSs.
Pasapera, A. M., Schneider, I. C., Rericha, E., Schlaepfer, D. D. & Waterman, C. M. Myosin II activity regulates vinculin recruitment to focal adhesions through FAK-mediated paxillin phosphorylation. J. Cell Biol. 188, 877–890 (2010).
Margadant, F. et al. Mechanotransduction in vivo by repeated talin stretch-relaxation events depends upon vinculin. PLoS Biol. 9, e1001223 (2011).
Smith, S. J. & McCann, R. O. A. C-terminal dimerization motif is required for focal adhesion targeting of talin1 and the interaction of the talin1 I/LWEQ module with F-actin. Biochemistry 46, 10886–10898 (2007).
Ellis, S. J., Pines, M., Fairchild, M. J. & Tanentzapf, G. In vivo functional analysis reveals specific roles for the integrin-binding sites of talin. J. Cell Sci. 124, 1844–1856 (2011).
Parsons, M., Messent, A. J., Humphries, J. D., Deakin, N. O. & Humphries, M. J. Quantification of integrin receptor agonism by fluorescence lifetime imaging. J. Cell Sci. 121, 265–271 (2008).
Kanchanawong, P. et al. Nanoscale architecture of integrin-based cell adhesions. Nature 468, 580–584 (2010).
Zhang, F., Saha, S. & Kashina, A. Arginylation-dependent regulation of a proteolytic product of talin is essential for cell–cell adhesion. J. Cell Biol. 197, 819–836 (2012).
Debrand, E. et al. Talin 2 is a large and complex gene encoding multiple transcripts and protein isoforms. FEBS J. 276, 1610–1628 (2009).
Franco, S. J. et al. Calpain-mediated proteolysis of talin regulates adhesion dynamics. Nature Cell Biol. 6, 977–983 (2004).
Huang, C. et al. Talin phosphorylation by Cdk5 regulates Smurf1-mediated talin head ubiquitylation and cell migration. Nature Cell Biol. 11, 624–630 (2009).
Bridgewater, R. E., Norman, J. C. & Caswell, P. T. Integrin trafficking at a glance. J. Cell Sci. 125, 3695–3701 (2012).
Albiges-Rizo, C., Frachet, P. & Block, M. R. Down regulation of talin alters cell adhesion and the processing of the a5b1 integrin. J. Cell Sci. 108, 3317–3329 (1995).
Martel, V. et al. Talin controls the exit of the integrin a5b1 from an early compartment of the secretory pathway. J. Cell Sci. 113, 1951–1961 (2000).
Priddle, H. et al. Disruption of the talin gene compromises focal adhesion assembly in undifferentiated but not differentiated ES cells. J. Cell Biol. 142, 1121–1133 (1998).
Liu, J. et al. Talin1 regulates integrin turnover to promote embryonic epithelial morphogenesis. Mol. Cell. Biol. 31, 3366–3377 (2011).
Margadant, C., Kreft, M., de Groot, D. J., Norman, J. C. & Sonnenberg, A. Distinct roles of talin and kindlin in regulating integrin α5β1 function and trafficking. Curr. Biol. 22, 1554–1563 (2012).
Bottcher, R. T. et al. Sorting nexin 17 prevents lysosomal degradation of β1 integrins by binding to the β1-integrin tail. Nature Cell Biol. 14, 584–592 (2012).
Steinberg, F., Heesom, K. J., Bass, M. D. & Cullen, P. J. SNX17 protects integrins from degradation by sorting between lysosomal and recycling pathways. J. Cell Biol. 197, 219–230 (2012).
Choi, C. K., Zareno, J., Digman, M. A., Gratton, E. & Horwitz, A. R. Cross-correlated fluctuation analysis reveals phosphorylation-regulated paxillin–FAK complexes in nascent adhesions. Biophys. J. 100, 583–592 (2011).
Zhao, Y. et al. Regulation of cell adhesion and migration by Kindlin-3 cleavage by calpain. J. Biol. Chem. 287, 40012–40020 (2012).
Bledzka, K. et al. Tyrosine phosphorylation of integrin β3 regulates kindlin-2 binding and integrin activation. J. Biol. Chem. 285, 30370–30374 (2010).
Qadota, H., Moerman, D. G. & Benian, G. M. A molecular mechanism for the requirement of PAT-4 (integrin-linked kinase (ILK)) for the localization of UNC-112 (Kindlin) to integrin adhesion sites. J. Biol. Chem. 287, 28537–28551 (2012).
Debrand, E. et al. Mice carrying a complete deletion of the talin2 coding sequence are viable and fertile. Biochem. Biophys. Res. Commun. 426, 190–195 (2012).
Desiniotis, A. & Kyprianou, N. Significance of talin in cancer progression and metastasis. Int. Rev. Cell Mol. Biol. 289, 117–147 (2011).
Kato, H. et al. The primacy of β1 integrin activation in the metastatic cascade. PLoS ONE 7, e46576 (2012).
Hynes, R. O. Integrins: bidirectional, allosteric signaling machines. Cell 110, 673–687 (2002).
Dong, X. et al. αVβ3 integrin crystal structures and their functional implications. Biochemistry 51, 8814–8828 (2012).
Bouaouina, M. et al. Zasp regulates integrin activation. J. Cell Sci. 125, 5647–5657 (2012).
Monkley, S. J. et al. Disruption of the talin gene arrests mouse development at the gastrulation stage. Dev. Dyn. 219, 560–574 (2000).
Wang, Y. et al. Loss of PIP5KIγ, unlike other PIP5KI isoforms, impairs the integrity of the membrane cytoskeleton in murine megakaryocytes. J. Clin. Invest. 118, 812–819 (2008).
Wernimont, S. A. et al. Contact-dependent T cell activation and T cell stopping require Talin1. J. Immunol. 187, 6256–6267 (2011).
Manevich-Mendelson, E. et al. Talin1 is required for integrin-dependent B lymphocyte homing to lymph nodes and the bone marrow but not for follicular B-cell maturation in the spleen. Blood 116, 5907–5918 (2010).
Lammermann, T. et al. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 453, 51–55 (2008).
Conti, F. J. et al. Progressive myopathy and defects in the maintenance of myotendinous junctions in mice that lack talin 1 in skeletal muscle. Development 135, 2043–2053 (2008).
Zou, W. et al. Talin1 and Rap1 are critical for osteoclast function. Mol. Cell. Biol. 33, 830–844 (2012).
Manso, A. et al. Talin1 has unique expression versus talin 2 in the heart and modifies the hypertrophic response to pressure overload. J. Biol. Chem. 288, 4252–4264 (2013).
Conti, F. J., Monkley, S. J., Wood, M. R., Critchley, D. R. & Muller, U. Talin 1 and 2 are required for myoblast fusion, sarcomere assembly and the maintenance of myotendinous junctions. Development 136, 3597–3606 (2009).
Bouvard, D., Pouwels, J., De Franceschi, N. & Ivaska, J. Integrin inactivators: balancing cellular fiunctions in vitro and in vivo. Nature Rev. Mol. Cell Biol. 14, 432–444 (2013).
Acknowledgements
Work in the authors laboratories was supported by the National Institutes of Health (NIH), the Medical Research Council, the Wellcome Trust, Cancer Research UK and the Medical Research Council. They thank members of the Calderwood laboratory for their input during preparation of the mansucript, B. Goult for help in preparing figure 1 and A. Kalli for contributions to figure 2c. They apologize to colleagues whose work could not be cited or fully discussed due to space limitations.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Related links
FURTHER INFORMATION
Glossary
- FERM domain
-
Found in various cytoskeletal-associated proteins, including band 4.1, ezrin, radixin and moesin, the proteins after which this domain was named. One role of this domain is to localize proteins to the plasma membrane. It typically contains three subdomains, F1, F2 and F3, that normally arranged in a cloverleaf formation.
- Nanodiscs
-
A synthetic model membrane system made from lipids and a scaffold protein. They are used to study incorporated membrane proteins, such as integrins, because they are relatively small, monodisperse and homogenous, yet provide a native-like environment.
- Leukocyte adhesion deficiency type III
-
(LADIII). A rare genetic disease characterized by severe bacterial infections and bleeding disorders. It is caused by mutations in the gene encoding kindlin 3 in haematopoietic cells.
- Multiscale molecular dynamics simulations
-
These numerically solve Newton's equations for interacting particles and follow their trajectories. They have become a powerful way of fine-tuning protein structure and dynamics. Major limitations are the relatively small size of the systems and the short timescales of the trajectories that can be studied. Multiscale simulations that combine simplified representations of molecules (coarse grain) with all atom representations (atomistic) circumvent some limitations.
- SAXS
-
(Small angle X-ray scattering). A technique that provides low-resolution information about the shape of objects. It depends on the analysis of the angular intensity distribution of X-rays scattered by molecules in solution.
- RAS-associating domain
-
(RA domain). A domain that is found in many effector proteins for the RAS family of small GTPases. The domain adopts a ubiquitin-like fold and supports binding of proteins containing an RA domain to 'active' GTP-loaded RAS family proteins.
- Pleckstrin homology domain
-
(PH domain). Domain that is found in a wide range of intracellular signalling proteins. It often binds to membranes via interactions with phosphatidylinositol-4,5-bisphosphate.
- Exocyst complex
-
An octameric protein complex that is involved in vesicle trafficking. It targets post-Golgi vesicles to the plasma membrane before vesicle fusion.
- PTB domain
-
(Phosphotyrosine-binding domain). Domain found in a wide range of intracellular signalling proteins. It commonly binds NPXY motifs; the Y may require phosphorylation to support binding.
- KRIT1
-
(Krev interaction trapped 1). This protein, which is encoded by the CCM1 gene, is a multidomain adaptor that is important for cell–cell and cell–matrix adhesion. Loss-of-function mutations in CCM1 cause predisposition to cerebral cavernous malformations, which are neurovascular anomalies that increase the risk of haemorrhagic stroke.
Rights and permissions
About this article
Cite this article
Calderwood, D., Campbell, I. & Critchley, D. Talins and kindlins: partners in integrin-mediated adhesion. Nat Rev Mol Cell Biol 14, 503–517 (2013). https://doi.org/10.1038/nrm3624
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrm3624
This article is cited by
-
Integrinβ-1 in disorders and cancers: molecular mechanisms and therapeutic targets
Cell Communication and Signaling (2024)
-
Inactivation of kindlin-3 increases human melanoma aggressiveness through the collagen-activated tyrosine kinase receptor DDR1
Oncogene (2024)
-
The niche matters: origin, function and fate of CNS-associated macrophages during health and disease
Acta Neuropathologica (2024)
-
Anoikis resistance––protagonists of breast cancer cells survive and metastasize after ECM detachment
Cell Communication and Signaling (2023)
-
Structure, signal transduction, activation, and inhibition of integrin αIIbβ3
Thrombosis Journal (2023)
