



Abstract
Cells respond to changing nutrient environments by adjusting the abundance of surface nutrient transporters and receptors. This can be achieved by modulating ubiquitin-dependent endocytosis, which in part is regulated by the NEDD4 family of E3 ligases. Here we report novel regulation of Pub1, a fission yeast Schizosaccharomyces pombe member of the NEDD4-family of E3 ligases. We show that nitrogen stress inhibits Pub1 function, thereby increasing the abundance of the amino acid transporter Aat1 at the plasma membrane and enhancing sensitivity to the toxic arginine analogue canavanine. We show that TOR complex 2 (TORC2) signalling negatively regulates Pub1, thus TORC2 mutants under nutrient stress have decreased Aat1 at the plasma membrane and are resistant to canavanine. Inhibition of TORC2 signalling increases Pub1 phosphorylation, and this is dependent on Gsk3 activity. Addition of the Tor inhibitor Torin1 increases phosphorylation of Pub1 at serine 199 (S199) by 2.5-fold, and Pub1 protein levels in S199A phospho-ablated mutants are reduced. S199 is conserved in NEDD4 and is located immediately upstream of a WW domain required for protein interaction. Together, we describe how the major TORC2 nutrient-sensing signalling network regulates environmental control of Pub1 to modulate the abundance of nutrient transporters.
Introduction
In all eukaryotic cells the external environment regulates cell fate. Highly conserved Target Of Rapamycin (TOR) signalling plays a key role in this control by responding to environmental cues, including stress and nutritional availability. This is achieved through TOR control of a series of metabolic processes, cell growth, migration, division, and differentiation. TOR signalling is extremely sensitive to changes in the cellular nutrient environment, and it is well established that reduced cellular energy levels and changes in amino acid concentrations are actively sensed to modulate TOR pathway activity (Laplante & Sabatini, 2012).
Several nutrient acquisition pathways support TOR control of anabolic cell growth (Selwan et al, 2016). These include autophagy, which breaks down cellular components to generate nutrients for anabolism, uptake of nutrients via surface transporters and receptor-mediated uptake of macromolecular nutrients (MacGurn et al, 2011; Laplante & Sabatini, 2012; Kim & Guan, 2015; Rispal et al, 2015; Roelants et al, 2017). Cells respond to alterations in their nutrient environment by regulating the abundance of surface nutrient transporters and receptors, in part by controlling their ubiquitin-dependent endocytosis.
Reciprocal regulation of TOR and nutrient pathways has been established because nutrients activate TOR, whereas TOR activity promotes endocytosis and inhibits autophagy. The mechanisms of TOR’s inhibition of autophagy to promote rapid cell proliferation in high nutrient environments is well-established (Kim & Guan, 2015). However, the impact of TOR stimulated endocytosis on nutrient utilisation is complex. On the one hand, TOR-controlled enhancement of endocytosis removes ion, carbohydrate and amino acid transporters from the plasma membrane, and may also reduce the surface population of macromolecular nutrient receptors, altogether reducing nutrient uptake (Ghaddar et al, 2014; Piper et al, 2014). On the other hand, endocytosis is also vital for the uptake of macromolecular nutrients such as low density lipoprotein (May et al, 2003).
TOR signalling comprises two structurally and functionally distinct multi-protein complexes. TOR kinases form TORC1 and TORC2 (TOR Complex 1 and 2), which are defined by unique subunits that are highly conserved across species. In mammalian cells the protein Raptor defines (mTORC1), whereas Rictor is exclusive to mTORC2 (Laplante & Sabatini, 2012). In the fission yeast Schizosaccharomyces pombe model, the focus of this study, Mip1 is the functional homolog of Raptor in TORC1, whereas Ste20 (Rictor homolog) defines TORC2 (Alvarez & Moreno, 2006; Hayashi et al, 2007; Matsuo et al, 2007) and Gad8, an ortholog of human AKT and SGK, is a well-established substrate of TORC2 (Matsuo et al, 2003; Ikeda et al, 2008; Du et al, 2016).
Studies in both yeast and mammalian cells have established mechanisms of TOR-regulated endocytosis and have documented both TORC1 and TORC2 dependent regulation of specific endocytic cargo (MacGurn et al, 2011; Rispal et al, 2015; Gaubitz et al, 2016; Grahammer et al, 2017; Roelants et al, 2017). For example, in Saccharomyces cerevisiae, amino acid permeases such as Can1 are down-regulated under nutrient-replete conditions by their ubiquitination by Rsp5 (a NEDD4 family E3 ligase), a process requiring the Rsp5 adaptor Art1 (MacGurn et al, 2011). This process is conserved in mammalian cells (Erpapazoglou et al, 2014; Piper et al, 2014; Boase & Kumar, 2015; Rosario et al, 2016). Ubiquitinated Can1 is then recognized by endocytic ubiquitin adaptors such as Ede1/Eps15 and Ent1/epsin and incorporated into clathrin-coated vesicles. TORC1 stimulates Can1 uptake pathway via the Npr1-dependent phosphorylation of Art1 and Can1, thereby enhancing Can1 ubiquitination (MacGurn et al, 2011). In mammalian neurons NEDD4 is a key regulator of neurite growth (Boase & Kumar, 2015) and studies have identified NEDD4-1 mRNA as a prominent target of mTORC1 regulated translation (Hsia et al, 2014). Human NEDD4-1 and NEDD4-2 (NEDD4L) also regulate ubiquitin-mediated autophagy, an mTORC1 controlled process (Chen et al, 2019; Lee et al, 2020). In fission yeast, the abundance of the Aat1 and Cat1 amino acid permeases on the plasma membrane increases after nitrogen starvation, and a contribution of Pub1 (NEDD4 family of E3 ligases), Tsc1/2 (Laplante & Sabatini, 2012) (an upstream inhibitor of TORC1) and TORC1 pathway to this localization has been demonstrated by several laboratories (Nefsky & Beach, 1996; Karagiannis et al, 1999; Matsumoto et al, 2002; van Slegtenhorst et al, 2004; Aspuria & Tamanoi, 2008; Nakase et al, 2012, 2013; Nakashima et al, 2014).
Each member of the NEDD4 HECT E3 ubiquitin ligase family comprises an amino-terminal Ca2+-phospholipid binding domain (C2), WW domains for protein to protein interaction, and a carboxy-terminal HECT domain containing its ligase activity (Boase & Kumar, 2015; Manning & Kumar, 2018; Huang et al, 2019). In the absence of Ca2+ binding to the C2 domain, conformational changes auto-inhibits NEDD4, whereas phosphorylation of NEDD4 on S347 and S348 by CK1 leads to its ubiquitination and degradation (Boase & Kumar, 2015). NEDD4-2 can also exist in an inactive form because AKT1- and SGK1-mediated phosphorylation of S342 and S428 promotes 14-3-3 binding to block NEDD4-2’s interaction with its substrates. In contrast, AMPK and JNK phosphorylation at the carboxy terminus is required for its activation (Boase & Kumar, 2015).
NEDD4 is expressed in most mammalian tissues and regulates a number of key substrates. Therefore, not surprisingly, dysregulation of NEDD4 ligases gives rise to a variety of diseases including cancer, cystic fibrosis, respiratory distress, hypertension, kidney disease, nervous system dysregulation, and epilepsy (Boase & Kumar, 2015; Manning & Kumar, 2018). In summary, NEDD4 ligase activity is regulated at multiple levels, including translation, phosphorylation, binding to accessory proteins and control of protein turnover. Consequently, the molecular mechanisms of its regulation are complex and are not fully understood. In this study we used the fission yeast model system to gain further insights into the mechanisms responsible for regulating the activity of this key E3 ligase. We show that nitrogen stress inhibits Pub1 function. TOR complex 2 (TORC2) and Gad8 (AKT) signalling negatively regulates Pub1 through their control of Gsk3 activity. Phosphorylation of Pub1 at serine 199 (a site conserved in NEDD4) is increased after TORC2/AKT inhibition and therefore Gsk3 activation. In summary, we show that the major TORC2 nutrient-sensing signalling network regulates Pub1 to modulate the abundance of nutrient transporters.
Results
TOR complex 2 (TORC2) negatively regulates Pub1
The cellular response to nutrient starvation is, in part, to increase the abundance of surface transporters to facilitate greater uptake of nutrients from the environment. In budding yeast, when nutrients are plentiful TORC1 inhibits Npr1 kinase to allow Rsp5 ubiquitin–dependent endocytosis of transporters (MacGurn et al, 2011). However, upon nutrient starvation when TORC1 activity is inhibited, so is the ubiquitin-dependent endocytosis, leading to higher levels of transporters at the plasma membrane (MacGurn et al, 2011). We previously undertook a global quantitative fitness profiling study to identify genes whose loss altered cell fitness in response to nitrogen stress. Not surprisingly, deletion of Pub1 (an NEDD4 family E3 ligase and the homolog of budding yeast Rsp5) increased cell fitness in response to nutrient stress (Lie et al, 2018). This is presumably because cells were able to import higher levels of nutrients due to reduced ubiquitin-dependent endocytosis of nutrient transporters.
With the aim of increasing our understanding of how Pub1 itself is regulated by changes to the cellular nutrient environment we exposed wild-type cells to nitrogen stress, by changing the nitrogen source from good to poor (here we changed from ammonia to proline - EMM2 to EMMP). This resulted in a 60% decrease in Pub1 protein levels (Figs 1A and S1A demonstrates that the antibodies detect Pub1). Therefore, in response to nitrogen stress when ubiquitin-dependent endocytosis is inhibited, the Pub1 E3 ligase is down-regulated.
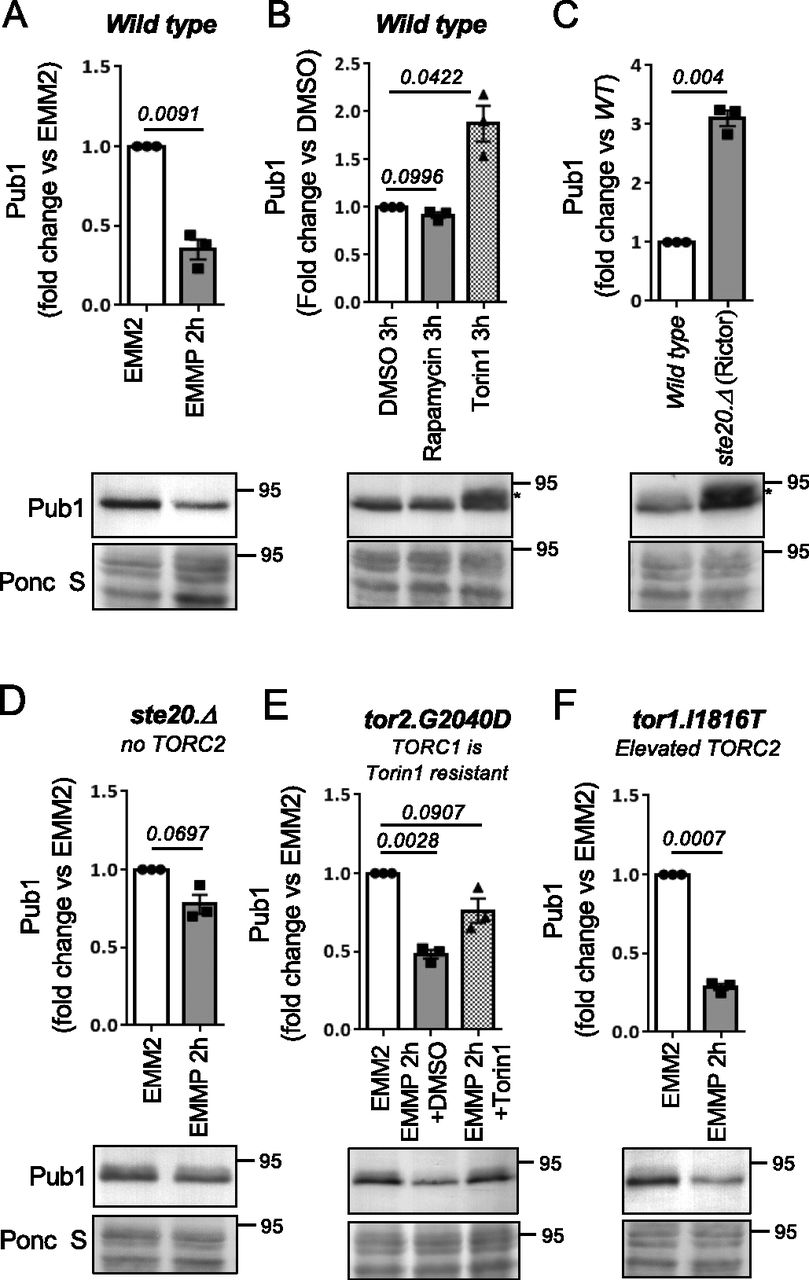
(A, B, C, D, E, F) Protein extracts were prepared from indicated yeast strains or treatments and immunoblotted for Pub1, Ponceau S is used to stain total protein. (B, C, E) After Torin1 treatment or inhibition of TORC2, a slower migrating form of Pub1 accumulates indicated by an asterisk (E). Torin1 was added at 15 μM to the EMMP medium of tor2.G2040D cells. Bars indicate fold change in levels versus indicated controls ± SEM, n = 3; n represents biologically independent experiments. Representative immunoblots are shown.

(A) Anti-Pub1 recognises Pub1. Protein extracts were prepared from indicated cells and immunoblotted for Pub1, Ponceau S is used to stain total protein. (B, C) Protein extracts were prepared from indicated yeast strain or treatments and immunoblotted for GFP to visualize Aat1.GFP, Ponceau S is used to stain total protein. (B) Bars mean fold change in levels versus EMM2 ± SEM, n = 3. n represents biologically independent experiments. Representative immunoblots are shown. (D) FM-4-64 does not interfere with Aat1.GFP fluorescence (compare with Fig 2A). Live cell imaging of Aat1.GFP in indicated strains. To differentiate between the two cell types (which were mixed at a ratio of 1:1 immediately before imaging), pub1.Δ cells were initially stained with FM-4-64 (indicated by a circle) for 45 min (accumulates in the vacuoles), before they were combined with unstained cells for immediate imaging. Fluorescence arbitrary units (AU) were determined as the brightest cell end/lowest nuclear background signal = (A/B).
To gain further insight into the environmental control of Pub1 protein levels, we treated wild-type cells grown in the good nitrogen source ammonia with the TOR kinase inhibitors rapamycin and Torin1 (ATP competitor TOR inhibitor 1 [Liu et al, 2012]) for 3 h. Rapamycin only inhibits TORC1, whereas Torin1 inhibits both TORC1 and TORC2 activities (Atkin et al, 2014). Rapamycin had no impact on Pub1 protein levels, whereas Torin1 promoted an increase in Pub1 levels (Fig 1B). At first glance this result appears contradicting. However, the imposition of nitrogen-stress to inhibit TORC1 has the opposite impact on TORC2 signalling, as previous reports demonstrated that TORC2 signalling is activated by nitrogen-stress in both fission yeast (after 1 h of nitrogen withdrawal) and human cells (Hatano et al, 2015; Laboucarié et al, 2017; Martín et al, 2017; Kazyken et al, 2019). Because rapamycin had no impact on Pub1 levels, TORC1 is unlikely to have a major role in regulating Pub1 levels. Our data therefore suggest that it is the inhibition of TORC2 by Torin1 that results in increased Pub1 protein levels (Fig 1B), whereas upon nitrogen stress when TORC2 signalling is activated Pub1 levels decrease (Fig 1A). In agreement with this notion, deletion of the TORC2-specific component ste20 (Rictor) also increased levels of Pub1 (Fig 1C).
The impact that nitrogen stress has on Pub1 protein levels was strongly diminished relative to wild-type when blocking TORC2 signalling in ste20.Δ (Rictor) mutants (Fig 1A and D). This indicates that active TORC2 is required for the observed decrease in Pub1 protein levels after nitrogen-stress. To test this further, we took advantage of our mutant in which we can inhibit TORC2 without affecting TORC1. Fission yeast Tor2 is the main kinase in TORC1, and we previously identified the tor2.G2040D mutation, in which TORC1 is resistant to Torin1 (Atkin et al, 2014). When the tor2.G2040D mutant was nitrogen stressed and Torin1 was added simultaneously (to inhibit only TORC2), the reduction of Pub1 due to media change to proline was diminished (Fig 1E). In contrast, in the TORC2 mutants Tor1.I1816T (Hálová et al, 2013), which has a small increase in TORC2 activity, Pub1 levels were reduced more efficiently upon nitrogen-stress (Fig 1A and F).
Together, our data suggest that TORC2 negatively regulates Pub1 and that environmental control of Pub1 protein levels after nitrogen stress is regulated by elevated TORC2 signalling. Note that after Torin1 treatment, a slower migrating form of Pub1 accumulates (indicated by an asterisk), indicating that TOR inhibition facilitates additional modification(s) of Pub1 (Fig 1C).
Aat1 amino acid transporter localization to the plasma membrane upon nitrogen stress requires TORC2 activity
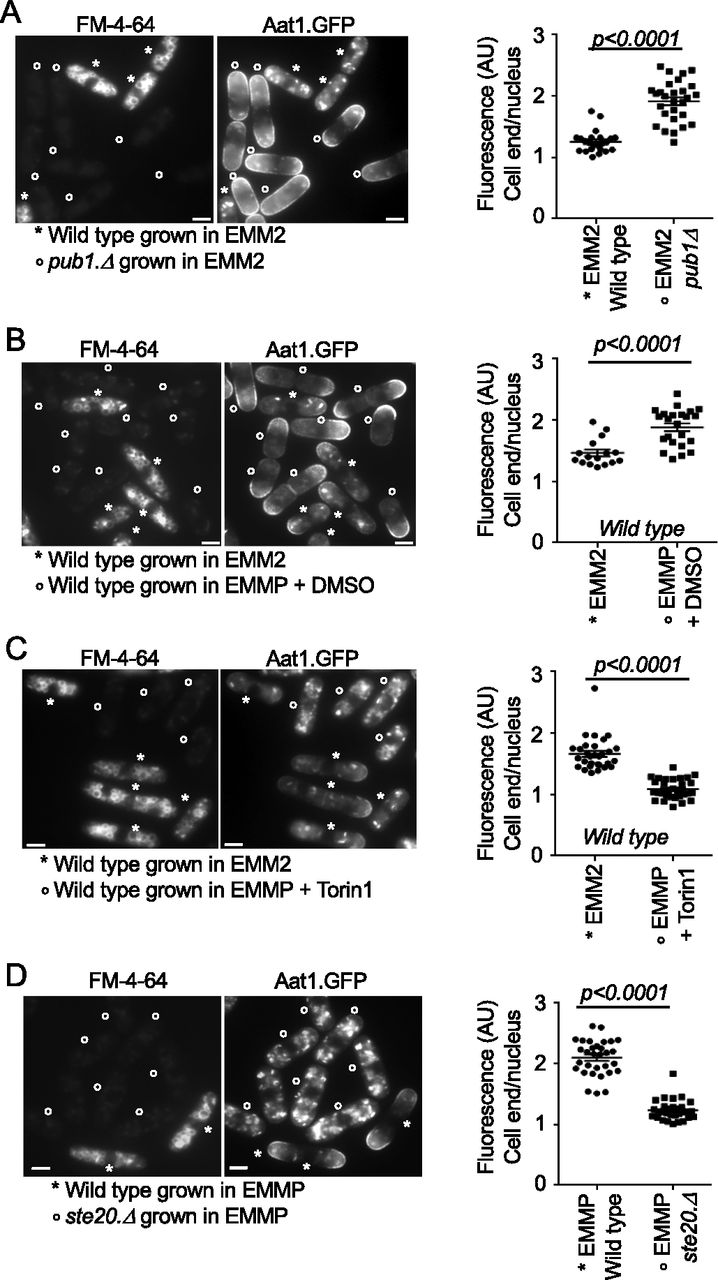
In fission yeast it is well-established that cells lacking Pub1 activity show increased abundance of the amino acid transporter Aat1 at the plasma membrane at cell tips (Matsumoto et al, 2002; Nakase et al, 2012). To visualize this, wild-type and pub1::ura4+ deletion cells were grown in EMM2, and wild-type cells were stained for 45 min with FM-4-64, which accumulates in the vacuoles, to differentiate between the two cell types when mixed 1:1 just before being imaged for Aat1.GFP localization (Fig 2A). Wild-type cells mainly had punctate cytoplasmic staining, previously attributed to localization at the Golgi (Liu et al, 2015). As expected, deletion of Pub1 increased Aat1 levels on the plasma membrane of cell ends by more than twofold, relative to background fluorescence seen in the nuclei (Fig 2A), without changing the levels of Aat1.GFP protein (Fig S1B and C). The same increase in Aat1.GFP localization in pub1 deletion strains was seen when the pub1.Δ cells were stained for 45 min with FM-4-64 before mixing with wild-type cells (Fig S1D). Thus, FM-4-64 does not interfere with Aat1.GFP localization or fluorescence.

(A, B, C, D) Live cell imaging of Aat1.GFP in indicated strains. To differentiate between the two cell types or treatments (which were mixed at a ratio 1:1), control cells were initially stained with FM-4-64 (accumulates in the vacuoles, indicated by a star) for 45 min, before they were mixed with unstained cells (indicated by circles) for immediate imaging. The relative fluorescence intensity of Aat1.GFP in all cells was quantified as: intensity at cell ends/cell tips (where cell growth occurs) relative to nuclear background fluorescence levels in the same cell (Fig S1D) (value = 1 represent identical arbitrary fluorescence intensity at the growing cell tips and in the nucleus of the same cell). FM-4-64 staining does not affect Aat1.GFP fluorescence (Fig S1D). Scale bar = 3 μm. All stats are calculated from images from one experiment. Representative images are shown. Similar results were obtained for three independent biological repeats. (A) Wild-type cells were strained with FM-4-64 (indicated by a star), to differentiate between the two cell types when mixed 1:1 with unstained pub1 deletion cells (indicated by a circle). (B, C) Wild-type cells grown in EMM2 were stained with FM-4-64. (B, C) Unstained cells (indicated by a circle) grown in EMM2 were treated before media shift for 1 h with DMSO (B). Torin1 at 25 μM (C). The cells were filtered into poor EMMP medium supplemented with Torin1 or DMSO, respectively, for a further 90 min before live cell imaging together with stained cells. (D) 90 min before imaging, wild type and ste20 deletion cells were shifted from EMM2 to EMMP medium. The wild-type cells were also stained with FM-4-64 (indicated by a star).
Imposition of nitrogen stress, by shifting wild-type cells from EMM2 into EMMP reduced Pub1 protein levels by 60% (Fig 1A). Consistent with such a reduction in Pub1, localization of Aat1.GFP at the plasma membrane was increased in nitrogen stressed cells (Fig 2B), whereas total Aat1.GFP protein levels remained unchanged (Fig S1B). However, the addition of Torin1 upon nitrogen stress to inhibit TORC2 signalling (Atkin et al, 2014) and increase Pub1 protein levels (Fig 1B and C) abolished Aat1.GFP localization to the plasma membrane (Liu et al, 2015) (Fig 2C). Finally, nitrogen-stress of the ste20 (Rictor) deletion, which block TORC2 function and thus increase Pub1 levels (Fig 1C) also blocked Aat1.GFP localization at the plasma membrane (Fig 2D). In summary, Aat1.GFP localization at the plasma membrane in poor nitrogen environments correlates with TORC2 regulation of Pub1 levels.
TORC2 and Gad8 (AKT/SGK) are required for Pub1 dependent nutrient uptake
TORC2 regulates Pub1 protein levels and the abundance of amino acid transporters on the cell membrane (Figs 1 and 2). To gain further insight into the molecular mechanism of TORC2 control of Pub1 function, we assessed the levels of Pub1 proteins in cells deleted of the only known substrate of TORC2 in fission yeast—the Gad8 kinase (AKT/SGK homolog) (Matsuo et al, 2003; Ikeda et al, 2008; Du et al, 2016). Like cells deleted for the TORC2 specific component ste20 (Rictor) (Fig 1C), elevated Pub1 levels were also observed in cells deleted of gad8 (Figs 3A and S2A). In the TORC2 mutants Tor1.I1816T, which has a small increase in TORC2 activity (Hálová et al, 2013), Pub1 levels appeared slightly reduced although this was not significant. Furthermore, blocking TORC2 signalling in gad8.Δ mutants only reduced Pub1 protein levels by ∼20% when nitrogen stressed (Fig 3B) relative to a 60% reduction in wild-type cells (Figs 1A and S2A), indicating that active Gad8 (AKT) is required to down-regulate pub1 after nitrogen stress.

(A, B) Elevated Pub1 in mutants of Gad8 (AKT). Protein extracts were prepared from indicated yeast strains and immunoblotted for Pub1, Ponceau S is used to stain total protein. Bars indicate fold change in levels versus indicated control ± SEM, n = 3. n represents biologically independent experiments. (C) Growth characteristics of indicated strains on EMM2 and EMM2 + 6 μg/ml canavanine and on EMMP or EMMP + 6 μg/ml canavanine. For all growth assays, similar results were obtained for three independent biological repeats.

(A) Protein extracts were prepared from indicated yeast strain or treatments and immunoblotted for Pub1, Ponceau S is used to stain total protein. (B) Growth characteristics of indicated strains on EMM2 and EMM2 + 20 μg/ml thiamine to repress promoter. Similar results were obtained for three independent biological repeats. (C) Protein extracts were prepared from indicated yeast strains and treatments and immunoblotted for Pub1, Ponceau S is used to stain total protein. Bars indicate fold change in levels versus EMM2 ± SEM, n = 3. n represents biologically independent experiments. Representative immunoblots are shown.
As shown above, cells increase the abundance of surface transporters to facilitate greater uptake of nutrients from the environment when stressed for nutrients (Fig 2B). We next used a simple well-established colony-forming growth assay to assess the role of TORC2 signalling on Pub1 function in vivo. Transport of canavanine into cells, a toxic arginine analogue, is in part regulated by the amino acid transporter Can1 (Fantes & Creanor, 1984), as the can1.1 canavanine-resistant mutant allele, in contrast to a wild-type strain, is able to form colonies when spotted from a serial dilution on agar-plates supplemented with canavanine. This is because the faulty transporter reduces the uptake of the toxic compound (Fig 3C) (Fantes & Creanor, 1984). In contrast, it is well-established that cells deleted for the Pub1 E3-ligase (which independently have reduced growth rate, even on EMM2 control media) are hypersensitive to canavanine (Fig 3C). This is because the block to ubiquitin-dependent endocytosis increases Can1 transporter abundance and therefore canavanine uptake (Fantes & Creanor, 1984; Aspuria & Tamanoi, 2008). Interestingly, can1.1 resistance is reduced in poor nutrient environments (EMM2 versus EMMP) (Fig 3C), which is consistent with decreased Pub1 function in EMMP (Fig 1A) and therefore increased transporter levels on the plasma membrane. This finding suggests that additional transporters may transport canavanine in the absence of Can1 function in the can1.1 mutant.
We next tested whether TORC2 and Gad8 control of Pub1 protein levels affected cells’ sensitivity to canavanine. Cells deleted of ste20 and gad8 display a substantially impaired growth rate compared with wild-type cells (data not shown), so are not ideal candidates to assess growth rates in our “canavanine-sensitivity” assay. We therefore took advantage of two other mutant strains to assess the consequences of increased or decreased TORC2/Gad8 activity. We previously showed that whereas a Gad8.T6D mutant (which reduces Gad8 function, through reduced TORC2 binding to Gad8) has normal growth rates on EMM2 media (Fig 3C), Gad8 activity is reduced albeit not blocked (Du et al, 2016). Reduced Gad8 activity in Gad8.T6D cells resulted in somewhat larger colony size (increased cell proliferation) on canavanine plates when compared with wild-type cells (Fig 3C), indicating that Pub1 function was modestly increased in this mutant to reduce the uptake of toxic canavanine. Importantly, Pub1 levels were also slightly increased in Gad8.T6D cells (Fig 3B), consistent with the modest increase in growth rate on canavanine plates. Notably, the opposite impact on growth rates was observed in cells with enhanced TORC2 activity in the Tor1.I1816T mutant (Hálová et al, 2013) (Fig 3C), as this mutant was sensitive to canavanine and exhibited a slight reduction in Pub1 protein levels. Together, these observations indicate that TORC2 and its downstream substrate Gad8 negatively impact on Pub1 protein levels and therefore regulate the levels of transporters on the membrane, which can transport the toxic arginine analogue canavanine into cells.
The TORC2 signalling pathway control Pub1 via Gsk3
To gain further insight into the molecular mechanism of TORC2 and Gad8 (AKT) control of Pub1 function we next considered Gsk3, as previous studies in human cells and fission yeast have shown that Gsk3 is a substrate of Gad8 (AKT) (Medina & Wandosell, 2011; Candiracci et al, 2019) and in fission yeast the TORC2 pathway was shown to regulate nutrient-dependent transcriptional elongation, through its inhibition of Gsk3 (Medina & Wandosell, 2011; Candiracci et al, 2019). Deletion of gsk3 decreased levels of Pub1 by 50% relative to wild-type cells (Fig 4A). Fission yeast Gsk31 is an ortholog of Gsk3. Whereas Pub1 levels remained unaffected in the gsk31.Δ deletion strain, Pub1 protein levels were further reduced in gsk3.Δ gsk31.Δ double deletion when compared with gsk3.Δ (Fig 4A), indicating that the two Gsk3 kinases are capable of functional redundancy upon deletion (Qingyun et al, 2016; Miao et al, 2020). Although the growth rate of the gsk3.Δ gsk31.Δ double mutant on minimal EMM2 medium is reduced (Fig 4B) it is sensitive to canavanine. Deletion of gsk3.Δ alone reduced colony size when exposed to the toxic compound, suggesting that Pub1 function was reduced in mutants lacking Gsk3 (Fig 4B). These observations fit with the reduced Pub1 protein levels seen in the gsk3.Δ mutant, which are further reduced in the gsk3.Δ gsk31.Δ double mutant (Fig 4A). We next asked whether the slow growth of gsk3.Δ gsk31.Δ double mutants (Fig 3B) could be rescued by Pub1 overexpression. Enhancing Pub1 levels only had a very minor enhancing impact on cell proliferation (Fig S2B). This is in line with the numerous cellular functions of Gsk3 in cells (Xu et al, 2009).
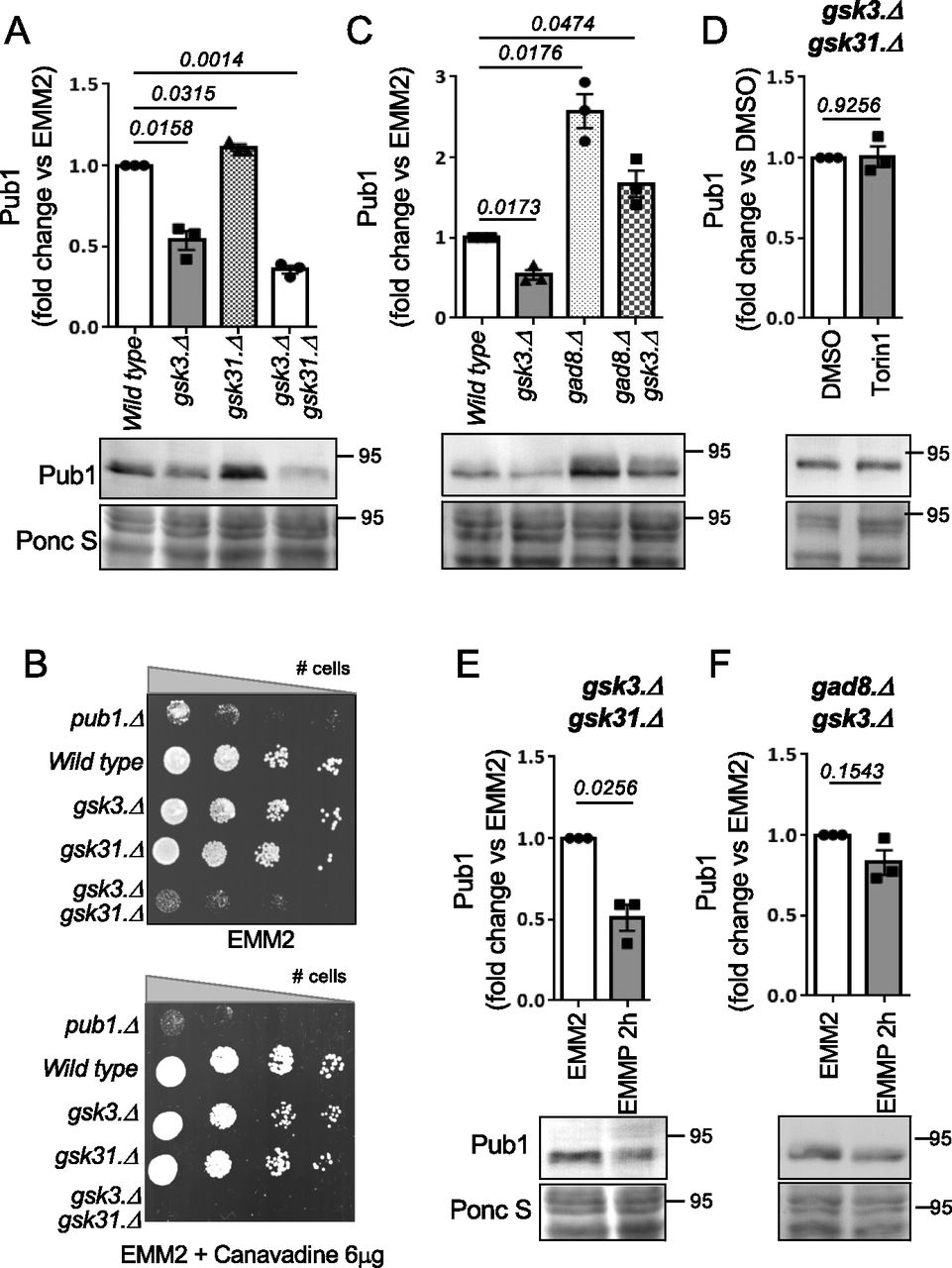
(A) Protein extracts were prepared from indicated yeast strains and immunoblotted for Pub1, Ponceau S is used to stain total protein. Bars indicate fold change in levels versus indicated control ± SEM, n = 3. n represents biologically independent experiments. Representative immunoblot is shown. (B) Growth characteristics of indicated strains on EMM2 and EMM2 + 6 μg/ml canavanine. For all growth assays, similar results were obtained for three independent biological repeats. (C, D, E, F) Protein extracts were prepared from indicated yeast strains and immunoblotted for Pub1, Ponceau S is used to stain total protein. Bars indicate fold change in levels versus indicated control ± SEM, n = 3. n represents biologically independent experiments. Representative immunoblots are shown.
We next analysed the reverse impact of Gad8 and Gsk3 on Pub1 levels further. If Gad8 regulates Pub1 through its demonstrated inhibition of Gsk3 activity, Pub1 levels in a double mutant are likely to resemble the levels seen in the gsk3.Δ mutant. The protein levels of Pub1 in the gsk3.Δ gad8.Δ double mutant were approximately half of those seen in gad8.Δ cells, and more than double that in gsk3.Δ cells (Fig 4C). These findings suggest that either the two kinases regulate Pub1 through independent mechanisms, or alternatively that the Gsk31 kinase present in the gsk3.Δ gad8.Δ double mutant is hyperactivated, due to lack of gad8, leading to elevated Pub1 protein levels compared with those seen in gsk3.Δ. Unfortunately, we were unsuccessful in generating a gsk3.Δ gsk31.Δ gad8.Δ triple deletion mutant to measure Pub1 protein levels and thus test this possibility. Nonetheless, in contrast to the situation in wild-type cells (Fig 1B), chemically inhibiting TORC2 signalling and therefore Gad8 with Torin1 failed to increase Pub1 protein levels in the gsk3.Δ gsk31.Δ double mutant (Fig 4D). This observation implies that the increase in Pub1 protein levels seen in cells defective in TORC2 signalling (Figs 1B and C and 3A) is controlled by Gsk3 activation.
In response to nitrogen stress, the down-regulation of Pub1 protein levels requires active TORC2 and Gad8 (Figs 1D and E and 3B). Therefore, Gsk3 activity is predicted to be dispensable for this lowering of Pub1 protein levels after nitrogen-stress because increased TORC2 signalling under nitrogen stress would inhibit Gsk3 (Medina & Wandosell, 2011; Candiracci et al, 2019). Indeed, the reduction in Pub1 levels was maintained upon nitrogen-stress of single gsk3.Δ and gsk31.Δ mutants and the gsk3.Δ gsk31.Δ double mutant (Figs 4E and S2C). We next tested the possibility that Gsk3 overexpression blocks Pub1 down-regulation after nitrogen stress. No significant change to Pub1 protein levels was seen in cells overexpressing Gsk3 compared with vector control (Fig S3A). However, previous reports have failed to identify any strong phenotypes upon Gsk3 overexpression in fission yeast (Plyte et al, 1996; Qingyun et al, 2016) apart from rescue of cell growth in cells lacking AMPK activity at 37C, which was also observed here (Fig S3B). Therefore, active Gad8 in nitrogen stressed cells may prevent significant activation of overexpressed Gsk3. We therefore turned to the gsk3.Δ gad8.Δ double mutant, in which Gsk31 appears to be hyperactivated because of deletion of gad8 deletion, leading to elevated Pub1 protein levels compared with those seen in gsk3.Δ (Fig 4C). Nitrogen stress of these cells, which lack the Gad8 inhibitor of Gsk31, failed to reduce Pub1 levels (Fig 4F).

(A) Overexpression of Gsk3 rescue growth of cells lacking AMPK activity. Growth characteristics of indicated strains on EMM2 at 25 and 37 degree. Similar results were obtained for three independent biological repeats. (B) Protein extracts were prepared from indicated yeast strains and treatments and immunoblotted for Pub1, Ponceau S is used to stain total protein. Ratios indicate fold change in Pub1 levels versus EMM2.
In summary, our observations suggest that Gsk3 activity protects Pub1 and that the reverse impact of TORC2/Gad8 and Gsk3 on Pub1 levels comes about because of TORC2/AKT–mediated Gsk3 inhibition (Candiracci et al, 2019) in fission yeast. Thus, lack of TORC2 activity enhances Gsk3 activity and consequently increases Pub1 protein levels.
Gsk3 blocks Pub1 degradation by the proteasome
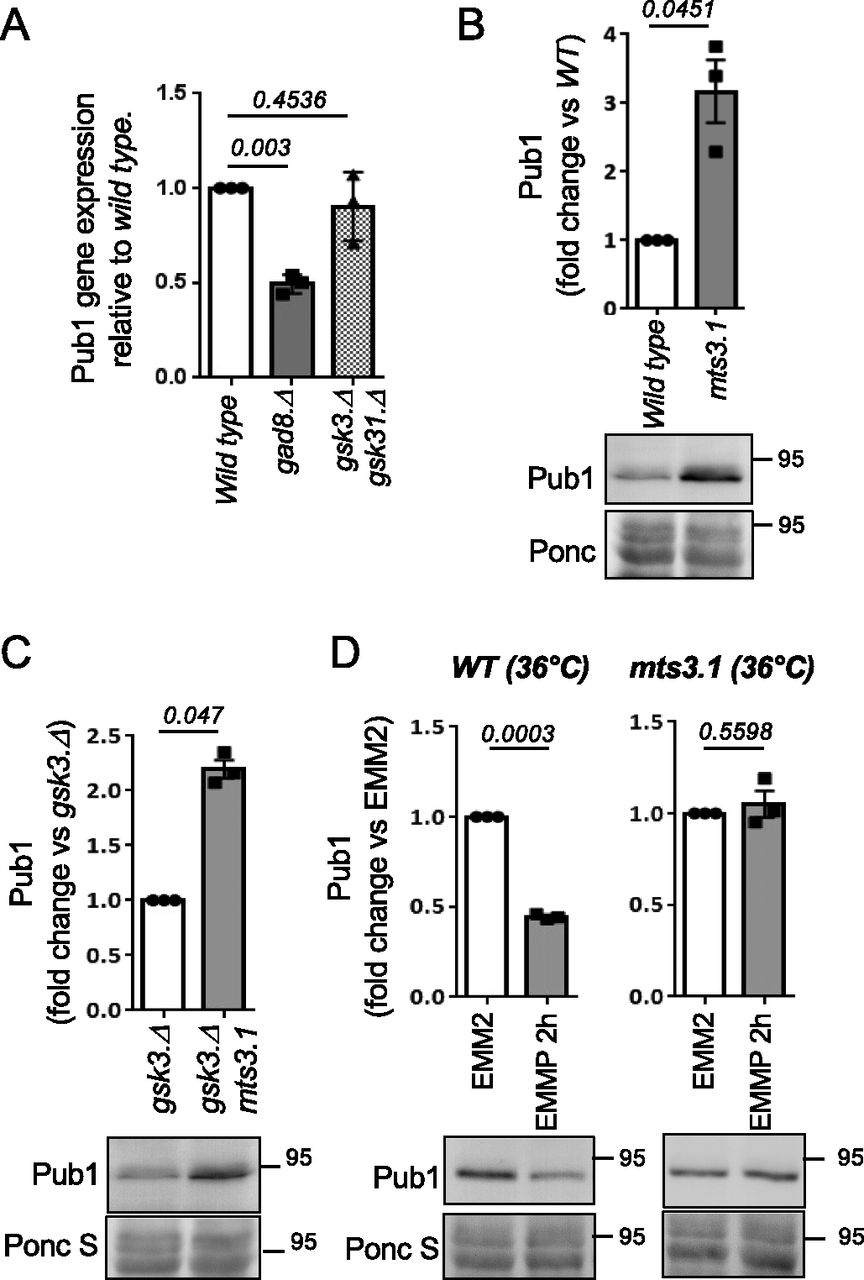
To further explore the mechanisms by which Gad8 and Gsk3 regulate Pub1 levels, we used quantitative PCR (qPCR) to assess the level of pub1 mRNA in the two kinase deletion strains. Interestingly, whereas the Pub1 protein levels are high in the gad8.Δ mutant, the mRNA levels are half that of wild-type cells, and the levels are unaffected in the gsk3.Δ gsk31.Δ mutants (Fig 5A). These observations suggest that the impact on protein levels in both mutants is independent of transcription. Auto-ubiquitination of S. cerevisiae Rsp5 and SCF-mediated degradation of human NEDD4 have been reported (Lam & Emili, 2013; Liu et al, 2014). Interestingly, a block to proteasome function in the mts3.1 proteasome mutant (Seeger et al, 1996) increased Pub1 levels threefold compared with wild type, to reach levels similar to that seen in the TORC2 mutant (Figs 5B and 1C). We conclude that Pub1 is degraded by the proteasome. Blocking proteasome function rescued Pub1 protein levels in the gsk3.Δ mutant (Fig 5C), demonstrating that Gsk3 activity is essential to prevent Pub1 degradation by the proteasome. Furthermore, lack of proteasome function in the mts3.1 mutant completely blocked Pub1 protein turnover (Fig 5D) after nitrogen stress, indicating that the proteasome is required for Pub1 destruction after under environmental nitrogen-stress.

(A) Levels of specific pub1 mRNA was measured by qPCR n = 3. Statistical significance was calculated. Unpaired t test with Welch’s correction (Prizm). n represents biologically independent experiments. (B, C, D) Protein extracts were prepared from indicated yeast strains and immunoblotted for Pub1, Ponceau S is used to stain total protein. Bars indicate fold change in levels versus indicated control ± SEM, n = 3. n represents biologically independent experiments. Representative immunoblots are shown. (D) Wild-type and mst3.1 cells were grown at 36°C for 6 h to inactive Mst3 before cell pelleting and protein extraction.
Phosphorylation of Pub1 serine 199 enhances protein levels after TORC2 inhibition
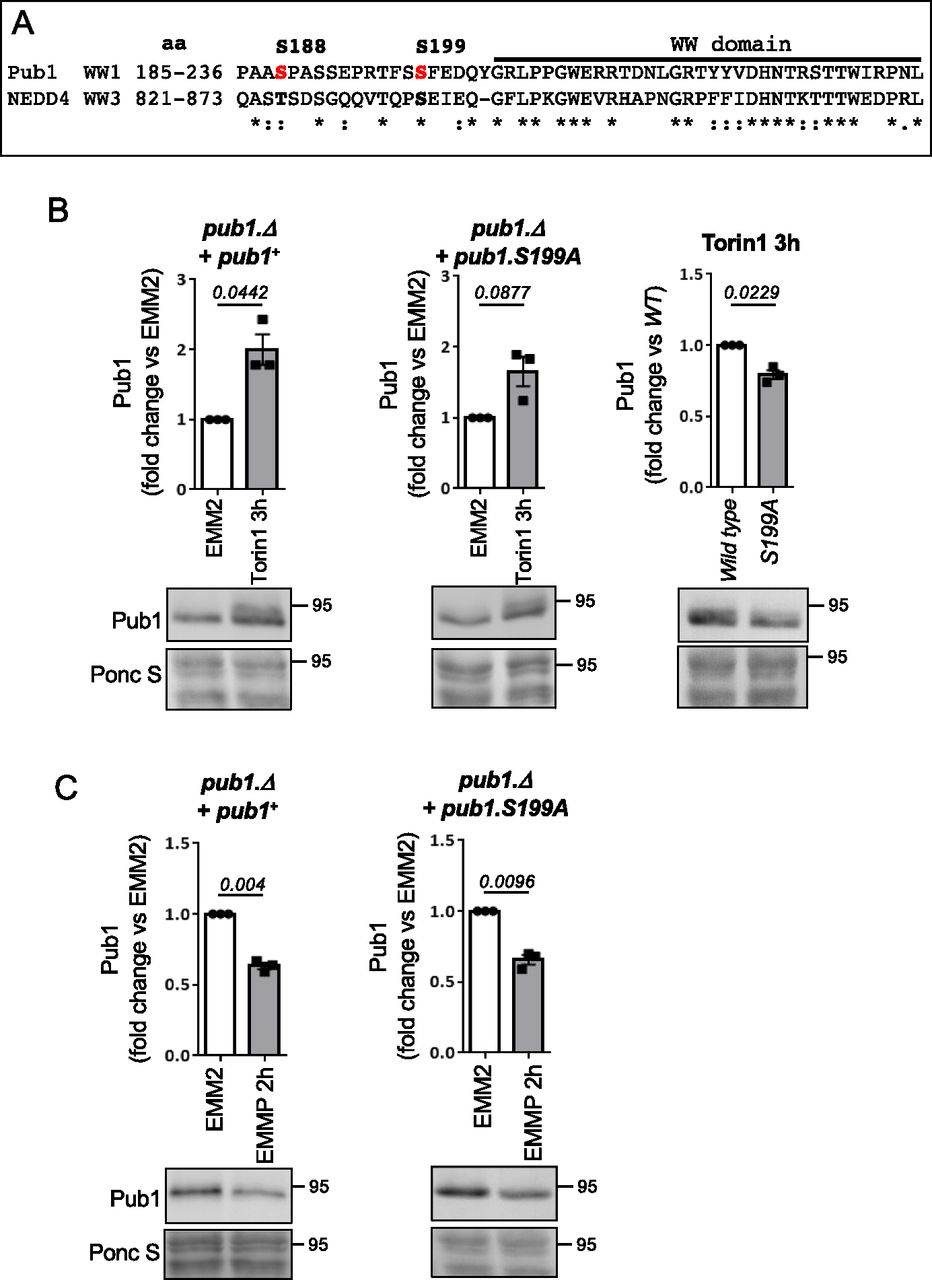
To increase our understanding of how Gsk3 blocks proteasome-mediated Pub1 degradation, we performed a quantitative, SILAC, and label-free mass spectrometry (MS)–based analysis (Humphrey et al, 2018) to identify Pub1 phosphorylation. Protein extracted from wild-type fission yeast that had been treated with Torin1 for 2 h to inhibit TORC2 and therefore activate Gsk3 was mixed 1:1 with either SILAC labelled or label free solvent treated controls. This identified five phosphorylation sites on Pub1 (Table S1). Interestingly, Pub1 serine 199 (S199) phosphorylation, which was reported previously in global screens but not characterised further (Kettenbach et al, 2015; Swaffer et al, 2018), was up-regulated 2.7-fold after Torin1 treatment. In contrast, upstream of S199, phosphorylation at serine 188 (S188) was decreased after Torin1 treatment (Fig S4A and Table S1). S199 is located directly upstream of the first WW domain in Pub1 and both S188 and S199 are conserved in human NEDD4 upstream of WW domain 3 (Serine 824 and Serine 835; Fig 6A). Phosphorylation of human NEDD4 S824 and S835 has not been reported. However, trypsin which is used routinely in shotgun proteomics studies generates a relatively long (48 amino-acids) NEDD4 peptide including these sites, which may not be identified in the MS analysis. We managed to generate phospho-specific antibodies to Pub1 S188 (Fig S4B and C). Consistent with our MS data, the relative levels of pS188 versus total Pub1 was reduced after 2 h of Torin1 treatment (Fig S4D). However, as pS188 is down regulated by Torin1, it is unlikely to represent a site phosphorylated by Gsk3.

(A) Chart illustrating changes to phosphorylation of Pub1 S188 and S199 after Torin1 treatment for 2 h by mass spectrometry (see Table S1). (B, C, D, F) Protein extracts were prepared from indicated yeast strains and treatments and immunoblotted for Pub1 or Pub1 pS188, Ponceau S is used to stain total protein. (B, C) Anti-Pub1 S188 recognise Pub1 and is phospho-specific. (C) Indicated Ponceau S–stained PVDF membranes were cut and incubated with or without lambda phosphatase for 1 h, to dephosphorylate proteins before immunoblotting. (D) 2 h of Torin1 treatment reduce phosphorylation of Pub1 serine 188. (F) Pub1 levels are unaffected by the status of serine 199 phosphorylation in good nutrient environments. (E) Sequence alignment of LRP6 (an established GSK3 substrate, which do not need a priming kinase), Pub1 S199, and human NEDD4 S835; also see sequence alignment in Fig 6A.

(A) Sequence alignment of Pub1 S188 and S199 (shown in red) with human NEDD4 homolog, aa indicates amino acids. (B, C) Protein extracts were prepared from indicated yeast strain and immunoblotted for Pub1, Ponceau S is used to stain total protein. Bars indicate fold change in levels versus EMM2 ± SEM, n = 3. n represents biologically independent experiments. Representative immunoblots are shown.
As described previously, in cells lacking Gsk3 activity Torin1 failed to increase Pub1 protein levels (Fig 4C) and failed to accumulate the slower migrating form of Pub1 (likely to represent the increase in phosphorylation) seen in wild-type cells (Fig 1B). Gsk3 commonly phosphorylates a primed sequence S/T-X-X-X-S/T(P) pre-phosphorylated by another kinase (Beurel et al, 2015). However, priming-independent GSK3 phosphorylation has also been reported in cells, for example, no priming kinase is required for LRP6 serine 1,572 phosphorylation by Gsk3 (Zeng et al, 2005, 2008; MacDonald et al, 2008). Interestingly, the sequence downstream of LRP6 S1572 is very similar to Pub1 S199 and NEDD4 835 (Fig S4E), hence S199 may be a direct Gsk3 site. To analyse the role of Pub1 S199 phosphorylation after Torin1 treatment, we mutated the serine to a phospho-blocking mutant alanine (A). A pub1 deletion strain was transformed with wild-type Pub1 and the S199A mutant. Torin1 was able to enhance pub1 levels in both wild-type and mutant S199A (Fig 6B); however, Pub1 levels in Pub1 S199A failed to accumulate to the level of wildtype (Fig 6B).
Whether Pub1 S199 is a direct Gsk3 site remains to be established; however, it is unlikely to be the only site on Pub1 regulated by Gsk3, as Pub1 still accumulated in the S199A mutant albeit not to the level of wild type. Pub1 levels in unstressed condition were unaffected in S199A mutants (Fig S4F). Furthermore, degradation after nitrogen stress was unaffected by the S199A mutation (Fig 6C), consistent with the notion that TORC2 signalling inhibits Gsk3 under nitrogen-stress, and hence pub1 is still degraded in Gsk3 null cells (Fig 4E).
Discussion
Here, we show for the first time that the fission yeast NEDD4 family of E3 ligase Pub1 is regulated by the nutrient environment and the major nutrient sensing TORC2 pathway, to control the levels of amino acid transporter on the plasma membrane and thus nutrient uptake. Previous studies have established mechanisms of both TORC1- and TORC2-dependent regulation of specific endocytic cargo and membrane transport in both yeast and mammalian cells (MacGurn et al, 2011; Rispal et al, 2015; Gaubitz et al, 2016; Grahammer et al, 2017; Roelants et al, 2017). However, we now show that TORC2 and its downstream substrate Gad8 (AKT) negatively regulates Pub1 function via Gsk3 summarised in Fig 7. We demonstrate that Gsk3 protects Pub1 from proteasomal degradation, as blocking proteasomal function in the mts3.1 mutant restores Pub1 protein levels in cells lacking Gsk3 activity (Fig 4D). Importantly, in both fission yeast and human cells, it is well established that AKT inhibits Gsk3 (Medina & Wandosell, 2011; Candiracci et al, 2019). Thus, the increase in Pub1 levels upon TOR inhibition with Torin1 is abolished in cells lacking Gsk3 activity (Fig 4D).

TORC2 and Gad8 signalling regulates Pub1 through demonstrated inhibition of Gsk3 activity. Thus, reduced TORC2 activity (Torin1 or gad8.T6D mutant cells) enhances Gsk3 activity and consequently increases Pub1 Function. In contrast, nitrogen stress known to activate TORC2 signalling (or tor1.T1816I mutant cells) inhibit Gsk3 activity to reduce Pub1 function.
When cells experience changes to their nutrient environment they respond by increasing the abundance of surface nutrient transporters, in part through down regulating their ubiquitin-dependent endocytosis. In agreement, we demonstrate TORC2-dependent Pub1 protein turnover through Gsk3 inhibition and proteasomal degradation after nitrogen stress (Figs 1D, 4F, and 5D). This, in turn results in increased Aat1 amino acid transporter abundance on the plasma membrane (Fig 2B) and increased sensitivity to the toxic arginine analogue canavanine (Fig 3C) summarised in Fig 7.
Our previous investigation of global quantitative fitness to detect genes whose deletion altered cell fitness in response to nitrogen stress or inhibition of TOR signalling identified Pub1 (Lie et al, 2018). Cells deleted of pub1 displayed enhanced cell fitness when nitrogen stressed, presumably because cells lacking pub1 are able to import higher levels of nutrients, due to reduced ubiquitin-dependent endocytosis of nutrient transporters. In this screen, the deletion of Gsk3 also enhanced the fitness of cells grown on minimal medium (P = 0.108) (Lie et al, 2018), which is consistent with our observation that Pub1 protein levels are reduced in the gsk3.Δ mutant (Fig 4A). Increased viability upon nitrogen starvation of cells deleted of gsk3 has also been reported in an independent genome wide screen (Sideri et al, 2015). The role of Gsk3 in protein stability is well established, though in contrast to the protective role of Gsk3 on Pub1 protein stability we describe here, Gsk3 is known to prime many substrates for proteasome degradation, with more than 25 substrates identified in human cells that are degraded in a Gsk3-dependent manner (Xu et al, 2009).
Whether Pub1 S199 is a direct substrate of Gsk3 remains to be established. Whereas Gsk3 commonly phosphorylates a primed sequence S/T-X-X-X-S/T(P) pre-phosphorylated by another kinase (Beurel et al, 2015), priming independent GSK3 phosphorylation has also been reported (Zeng et al, 2005, 2008; MacDonald et al, 2008). The sequence downstream of Pub1 Serine 199 is similar to that of an established Gsk3 substrate (Fig S4E), highlighting this site as a candidate. Future experiments will establish whether Pub1 is a direct Gsk3 substrate. Human NEDD4 is also degraded by the proteasome, as phosphorylation of NEDD4 on S347 and S348 by CK1 leads to SCF-mediated ubiquitination and degradation (Boase & Kumar, 2015). However, the SCF phospho degrons DSGXXS or T-P-P-X-S are not conserved in Pub1, and deletion of CK1 activity in fission yeast does not increase Pub1 levels (data not shown). How Gsk3 protects Pub1 from proteasomal degradation is currently unclear. However, Pub1 protein levels were reduced in the S199A mutant compared with wild type, when cells were treated with Torin1 (Fig 6B), suggesting phosphorylation of this site is important.
Pub1 S199 phosphorylation has no impact on protein levels in nutrient-rich environments (Fig S4F), therefore, considering the close proximity of Pub1 S199 and NEDD4 S835 to their WW domains (Fig 6A) phosphorylation is likely to regulate protein–protein interactions of adaptor proteins important for function under nutrient stress. Future experiments will address this.
Whether the regulation of Pub1 we report here is conserved in human cells is unclear at this stage. In human cells, GSK3 negatively regulates glucose homeostasis (Lee & Kim, 2007). Furthermore, insulin and growth factor signalling, which activates mTORC1, mTORC2, AKT, and S6K inhibit GSK3 activity (Medina & Wandosell, 2011) and thereby increase glycogen synthesis. In contrast, NEDD4 enhances insulin and growth factor signalling (Cao et al, 2008; Fukushima et al, 2015). Our observation suggests that reduced GSK3 activity as a result of insulin signalling may decrease NEDD4 function and thus put a brake on insulin and growth factor signalling through a negative feedback loop. However, in human cells, NEDD4 can directly bind to and ubiquitinate AKT which is prior phosphorylated on pS473, to degrade active AKT (Huang et al, 2020). Thus, decreased GSK3 activity and therefore reduced NEDD4 function would increase active AKT pS473, providing a positive feedback for insulin and growth factor signalling and glucose uptake to counteract the aforementioned negative feedback and thereby establish a steady state. Therefore, if conserved, the mechanism described here would most likely only impact hormone signalling and glucose uptake when this pathway is interacting with other signalling pathway(s) that alter the steady state.
In summary, here we show that Gsk3 protects Pub1 function, in part through S199 phosphorylation. We also provide the first evidence of NEDD4 family E3 ligase being regulated by nitrogen stress and TORC2 signalling to reduce ubiquitin-dependent endocytosis, thus increasing the abundance of amino acid transporters on the plasma membrane when nutrient levels are challenging.
Materials and Methods
Yeast cell cultures
Strains used in this study (Table 1). All cultures were grown at 28°C and cultured in log phase for 48 h. Cells were inoculated in in Edinburgh minimal medium (EMM2-N) (ForMedium) (Fantes, 1977) supplemented with NH4Cl (EMM2) (Petersen & Russell, 2016). Media change to (EMM2-N) (ForMedium) (Fantes, 1977) supplemented with proline (EMMP) (Petersen & Russell, 2016) was done by filtering cells, followed by resuspension into prewarmed EMMP.
Strains used in this study.
SILAC labeling and harvesting culture for mass spectrometry
SILAC labeling: Cells were inoculated in Edinburgh minimal media (EMM2-N) (ForMedium) (Fantes, 1977) supplemented with 20 mM L-Glutamic acid (EMMG) (Petersen & Russell, 2016) and 75 mg/l of either light (L-arginine monohydrochloride [Sigma-Aldrich] and L-lysine monohydrochloride [Sigma-Aldrich]) or medium (lysine-L, 2HCl 4.4.5.5-D4 [Cat code DLM-2640; Eurisotop], arginine-L, HCl, U-13C6 99%13C [Cat. no. CLM-2265; Eurisotop]) amino acids. Cells were cultured in log phase for 30 h to ensure complete incorporation of labelled amino acids into the proteome. Early log phase cultures at 3.5 × 106 cell/ml were treated with 15 µMTorin1 or DMSO control. Cells were collected by filtration (MF-Millipore Filter, 1.2 μm pore size Cat. no. RAWP04700; Millipore) washed with 15 ml TBS, resuspended in an appropriate volume of ice cold sterile ddH2O and dropped directly into liquid nitrogen to produce frozen cell droplets.
Mass spectrometry
SILAC mass spec analysis of samples processed using a SPEX Sample Prep LLC 6850 Freezer Mill in presence of liquid nitrogen, were performed as described previously (Humphrey et al, 2018). Data were analysed with MaxQuant (Cox & Mann, 2008) (v1.6.0.9) using the Andromeda search engine (Cox et al, 2011) to query a target-decoy database of S. pombe from UniProt (September 2019 release).
Drug treatment
L-Canavanine sulfate salt (Cat. no. C9758; Sigma-Aldrich) was added to EMM2 agar plates at a concentration of 6 μg/ml, Torin1 (Cat. no. 4247; TOCRIS) was used at a concentration of 15 and 25 μM. Rapamycin (Cat. no. R0395; Sigma-Aldrich) was used at a concentration of 300 ng/ml.
Generation of pub1.S199A mutant
The pub1 serine 199 point mutation was generated by site directed mutagenesis of Rep41-Pub1. Transformation of a pub1::ura4+ deletion and selection on plates lacking leucine (Petersen & Russell, 2016) were used to select transformants. Strains were grown in the presence of 10 μM thiamine to reduce the level of Pub1 expression.
Western blotting
TCA precipitation protocol was followed for S. pombe total protein extracts (Caspari et al, 2000). The following dilutions of antibodies were used in this study: 1/250 anti-Pub1 pS188 and 1/500 anti-Pub1 (custom made by Thermo Fisher Scientific, anti-Pub1 being non-specific to pS188) in PBS buffer, 1/500 anti-GFP (Cat. no. 11814460001; Roche) in TBS buffer, skim milk was used as blocking agent. Alkaline phosphatase coupled secondary antibodies were used for all blots followed by direct detection with NBT/BCIP (VWR) substrates on PVDF membranes.
GraphPad Prism 6.07 was used for data analysis. Unpaired t test with Welch’s correction (Prizm version 7) were used for all Western blots. 95% confidence of interval was used for calculating significance.
Fluorescent microscopy
Staining of vacuoles: SynaptoRed C2 (Equivalent to FM4-64) (Cat. no. 70021; Biotium) was added to the growth medium of cells (1 × 106 cells/ml) at a concentration of 1.5 μM for 45 min. Cultures of Stained and unstained cells were mix 1:1 and collected by filtration onto MF-Millipore Membrane Filter, 1.2 μm pore size (Cat. no. RAWP04700; Millipore). Cells were resuspended in the original growth media of the FM4-64–stained cells and subjected to live cell imaging immediately. Images of cells were obtained using a CoolSNAP HQ2 CCD camera. ImageJ was used to measure fluorescent intensities of Aat1.GFP. The relative fluorescence intensity (arbitrary units) of Aat1.GFP was quantified as: the highest intensity at the brightest cell end, relative to the lowest nuclear background fluorescence signal of the same cell (see Fig S1D), to allow for comparisons of separate images and experiments. Statistical significance was calculated using Unpaired t test.
RNA extraction and qPCR
RNA was extracted using TRIzol Reagent (Cat. no. 15596026; Thermo Fisher Scientific). In short, 1 × 107 cells in early log phase were collected by centrifugation. Cell pellets were snap-frozen in liquid nitrogen. 1 ml of Trizol and 200 μl of glass beads (Cat. no. 11079105; Biospec) were added to the cells. Cells were disrupted by a FastPrep-24 (MP) at 5 m/s for 60 s for three cycles in a cold room. Cell lysate was processed according to the manufacturer’s instructions. RNA pellets were resuspended in 50 μl of RNAse-free water. 1,000 ng of RNA subjected to DNA digestion by TURBO DNA-free Kit (Cat. no. AM1907).
First-strand cDNA were synthesized from 500 ng of RNA by using M-MLV Reverse Transcriptase, RNase H Minus, Point Mutant (Cat. no. M3683; Promega). DNAse-treated RNA, 500 ng Oligo (DT)15 (Cat. no. C1101; Promega) and 100 ng random hexamer (Cat. no. C1181; Promega) and heated to 70°C for 5 min, cooled to 4°C, and incubated on ice for 5 min. For reverse transcription, RNA, primers, dNTP mix (Cat. no. N0446S; Bio New England Lab), M-MLV RT (H-) Point Mutant were used. 1: 4 diluted first-strand cDNA were used for second strand synthesis of cDNA and qPCR using Power SYBR Green PCR Master Mix (Cat. no. 4367659; Thermo Fisher Scientific). Reactions were run in Rotor-Gene Q (QIAGEN) with initial activation at 95 for 10 min, followed by 40 cycles of 95°C for 15 s, 58°C for 1 min. Comparative Quantitation Analysis from Rotor-Gene Q series software produced Representative Takeoff vale from triplicates of each sample. 2−ΔΔCT method was used to calculate pub1 gene expression relative to housekeeping gene act1. Primers to amplify pub1 gene: Forward: CCCTTATTGGAATGAGACTTTTG; Reverse: GGGTCAACATTTCATCACCTC. Forward and Reverse Primers to amplify the control act1 gene was a described (Biswas et al, 2016).
Acknowledgements
We thank the Japanese National BioResource project for providing strains. This work was supported by the Australian Research Council (DP180101682) to SJ Humphrey and J Petersen, Flinders Foundation seeding grant, and Flinders Universities supported this work. P Woodman was supported by the Leverhulme Trust (RPG-2018-091).
Author Contributions
T Wang: investigation, methodology, and writing—review and editing.
P Woodman: conceptualization, data curation, funding acquisition, and writing—review and editing.
SJ Humphrey: conceptualization, formal analysis, funding acquisition, investigation, methodology, and writing—review and editing.
J Petersen: conceptualization, formal analysis, supervision, funding acquisition, project administration, and writing—original draft, review, and editing.
Conflict of Interest Statement
The authors declare that they have no conflict of interest.
- Received March 30, 2021.
- Revision received January 24, 2022.
- Accepted January 25, 2022.
- © 2022 Wang et al.
This article is available under a Creative Commons License (Attribution 4.0 International, as described at https://creativecommons.org/licenses/by/4.0/).
References




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
