




Article Figures & Data
Figures
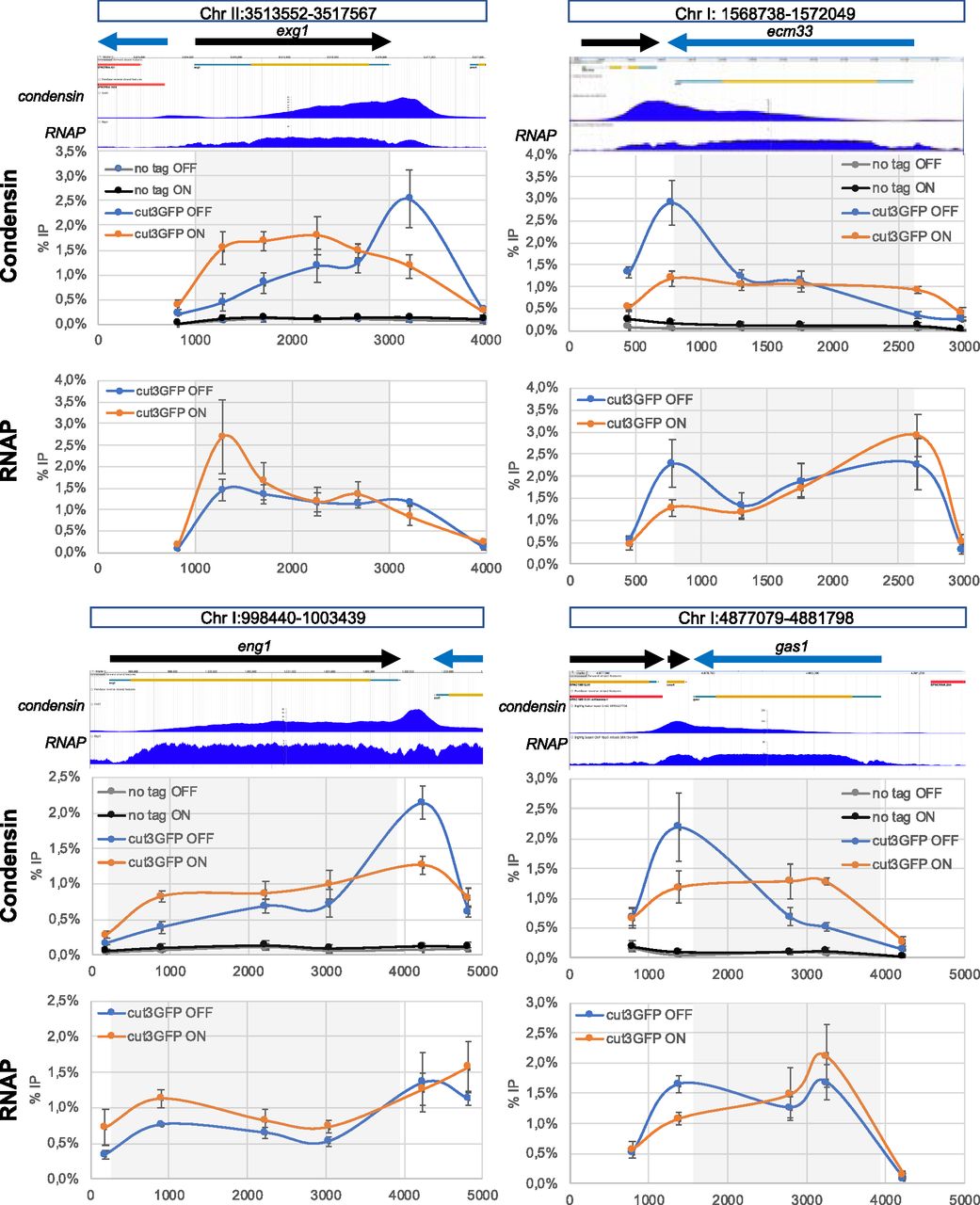
- Figure 1. Distribution of condensin and RNA polymerase 2 upon flipping of exg1.
(A) ChIP-seq profiles of condensin (left) and the RNA polymerase subunit Rpb5 (right) around exg1 in mitotic cells. The ChIP-seq data are indicated by their Sequence Read Archive numbers and were generated in Sutani et al (2015) and Kakui et al (2017), respectively. (B) Cells were synchronized in mitosis and ChIP-qPCR in two different biological replicates was used to determine the distribution of condensin (left) and Rpb1 (right) around exg1. (C) Same as (B) when the orientation of exg1 has been flipped over. The scheme above shows the organization of the chromosome around exg1 in the wild-type (top) and in the reversed (bottom) orientations. Vertical dotted lines indicate the region of the chromosome that has been flipped over. Grey squares indicate the position of the exg1 transcription unit. The % IP were normalized using the values given at the site within the gene body indicated by the red vertical dotted line (exg1#1). The raw data are shown in the source data files.
Source data are available for this figure.
Source Data for Figure 1[LSA-2021-01046_SdataF1.xlsx]
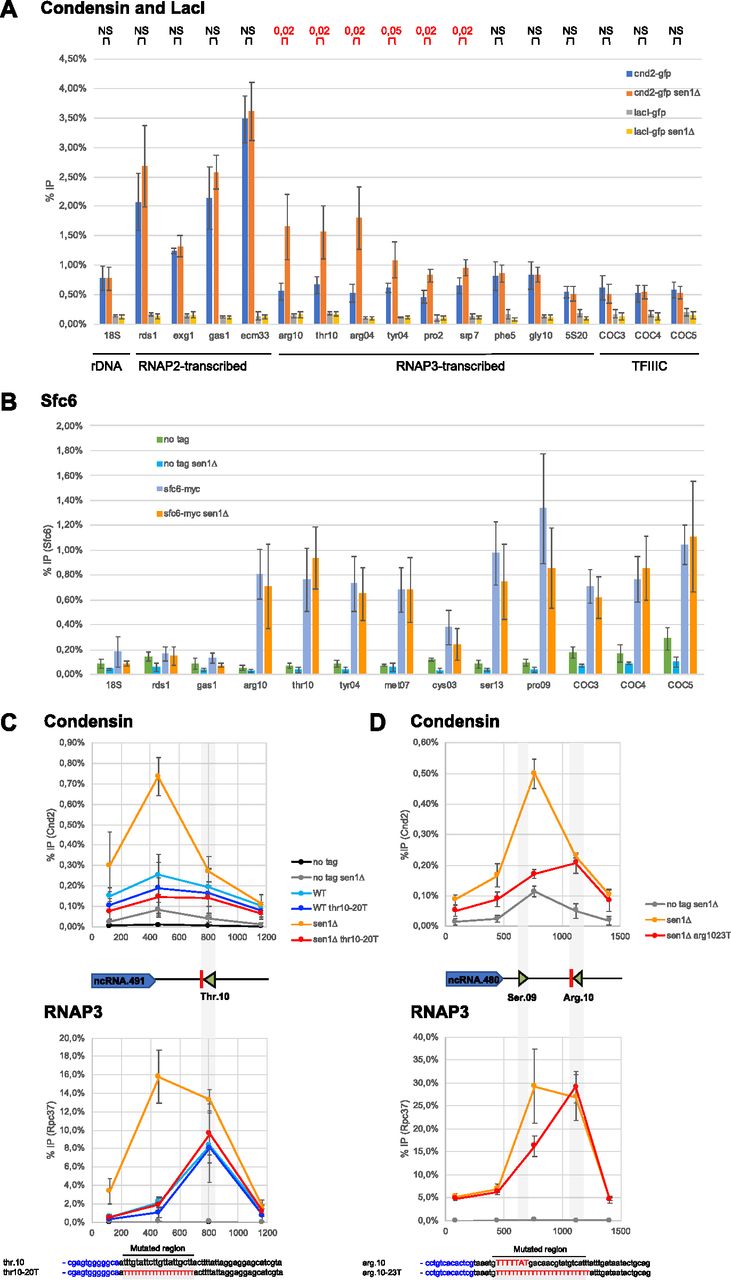
- Figure 2. RNA polymerase 3 transcription defects induced by lack of Sen1 trigger the accumulation of condensin.
(A) Cells were synchronized in metaphase and the association of condensin (Cnd2-GFP) or the heterologous LacI (lacI-GFP) at the indicated loci was investigated by ChIP-qPCR in the presence and in the absence of Sen1 (mean ± std of four biological replicates; P-values determined by the test of Wilcoxon Mann-Whitney are indicated above the graph). (B) The association of the TFIIIC component Sfc6 at the indicated loci was investigated by ChIP-qPCR in cells synchronized in metaphase (mean ± std of five biological replicates). (C, D) Distribution of condensin (cnd2-GFP, top) and RNA polymerase 3 (rpc37-flag, bottom) around SPCTRNATHR.10 (C) and SPCTRNAARG.10 (D) in mitotic cells, in the presence or not of super-terminator sequences (thr10-20T and arg10-23T, respectively) which correct the transcription termination defects in the absence of Sen1 (Rivosecchi et al, 2019) (compare the yellow and red curves). (C, D) Results are presented as (mean ± std) of three (C) or four (D) biological replicates.
Source data are available for this figure.
Source Data for Figure 2[LSA-2021-01046_SdataF2.xlsx]
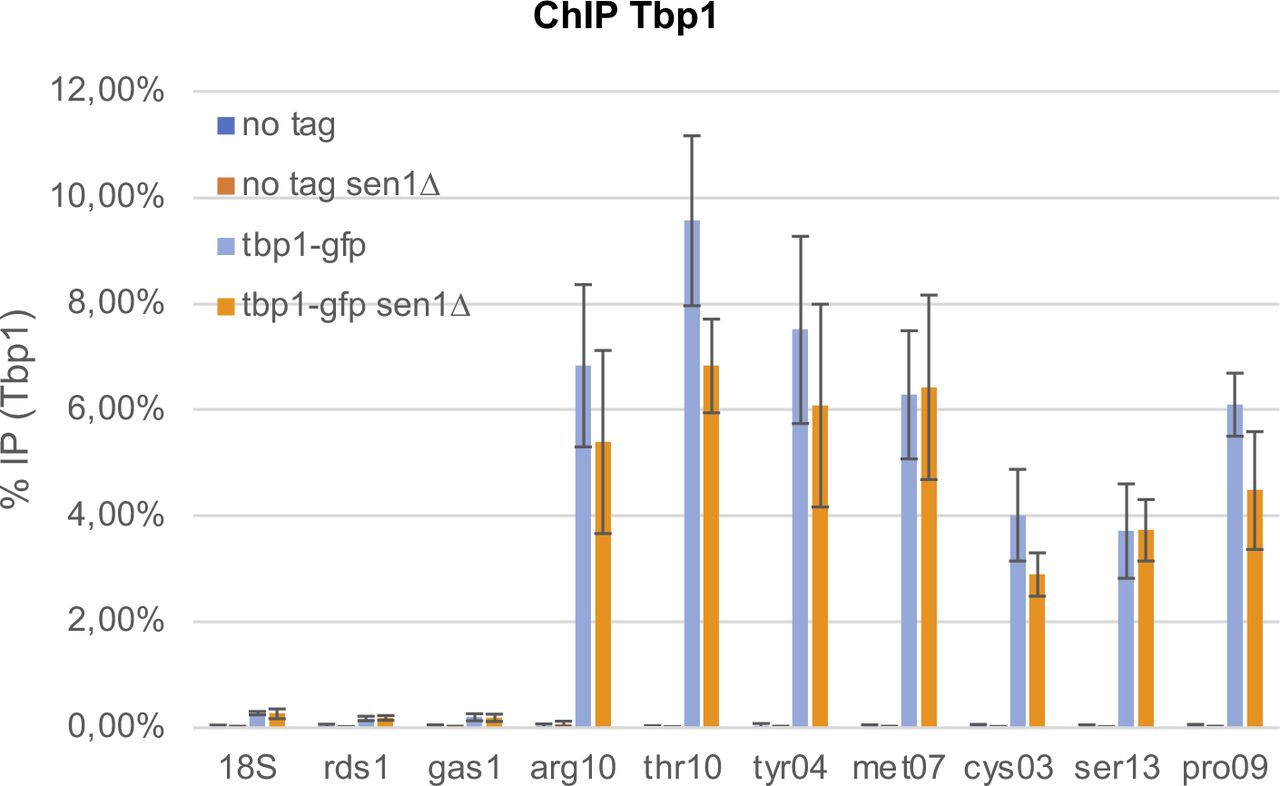
- Figure S1. Lack of Sen1 does not significantly alter the association of Tbp1 at RNA polymerase 3-transcribed genes.
Cells were synchronized in metaphase by depleting Slp1 (see the Materials and Methods section). The association of Tbp1 at the indicated loci was investigated by ChIP-qPCR (mean ± std of four biological replicates).
- Figure 3. The over-expression of tfs1DN alters significantly the distribution of condensin around RNA polymerase 2-transcribed genes.
Cells were synchronized in metaphase and the association of condensin (cnd2-GFP) at the indicated loci was investigated by ChIP-qPCR (mean ± std of three biological replicates). Cells carried a plasmid allowing the AhTET-induced over-expression of tfs1-DN, as described previously (Lemay et al, 2014). DMSO was used as control. For each locus investigated, the normal distribution of condensin and RNA polymerase 2 as determined by ChIP-seq is shown above, as published in Sutani et al (2015) and Kakui et al (2017), respectively. The raw data are shown in the source data files.
Source data are available for this figure.
Source Data for Figure 3[LSA-2021-01046_SdataF3.xlsx]
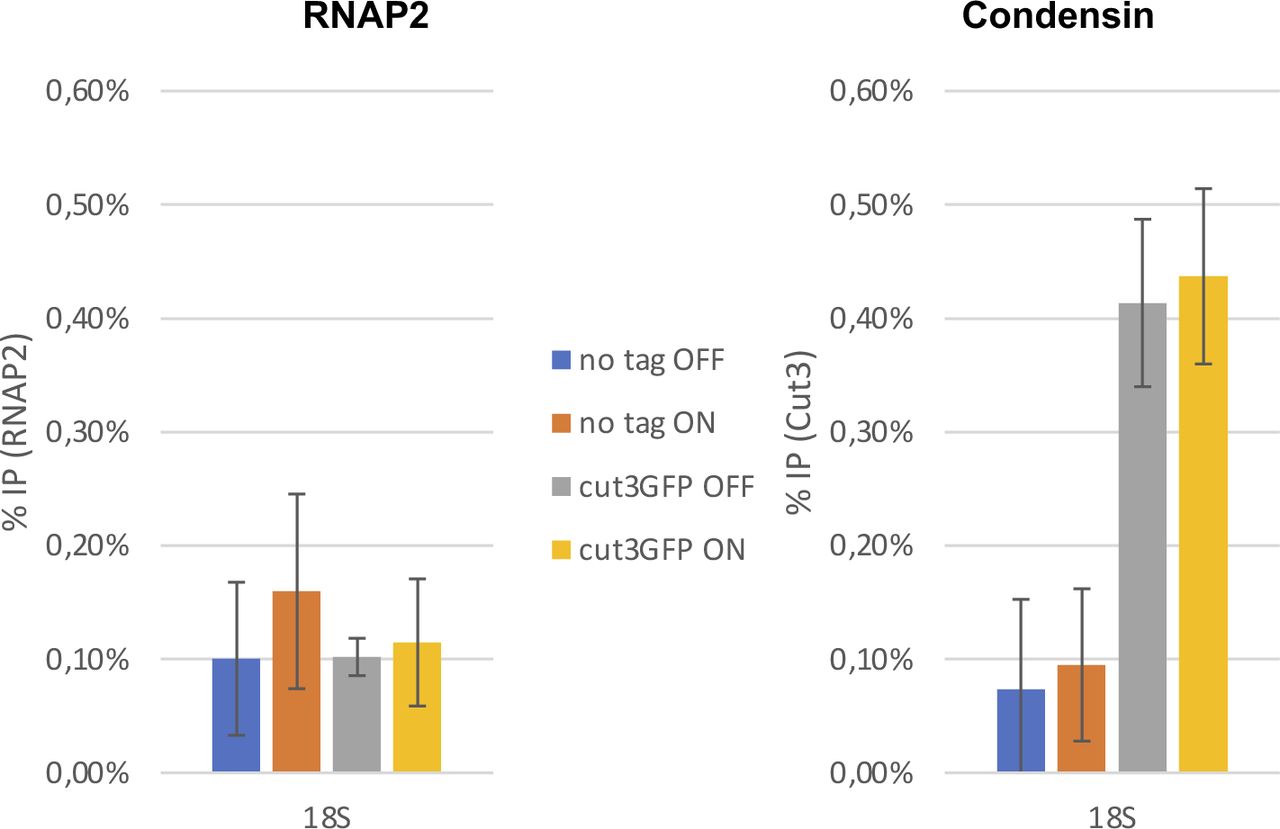
- Figure S2. The over-expression of tfs1DN does not interfere with the occupancy of condensin at the rDNA.
The samples described in Fig 3 were used to monitor the occupancy of condensin within the RNA polymerase 1-transcribed 18S transcription unit.
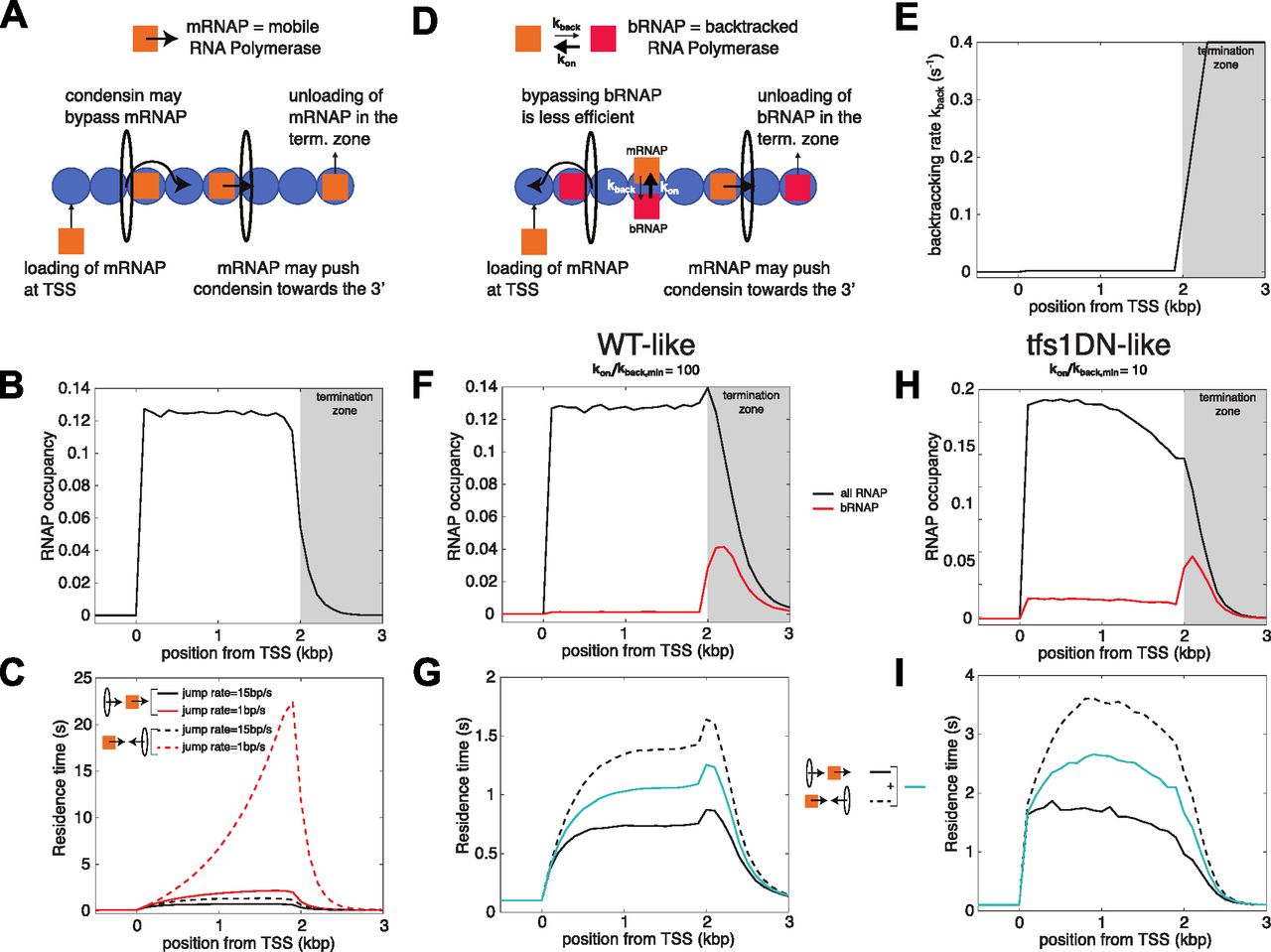
- Figure 4. Mathematical models formalize the role of RNA polymerase (RNAP) backtracking in the specific accumulation of condensin in the termination zone of active genes.
(A) Interplay between the translocation of condensin and transcription—simple model (Brandão et al, 2019). Condensin translocates along chromatin from either 5′ to 3′ or 3′ to 5′. RNAPs bind to TSS, translocate unidirectionally from 5′ to 3′ and unbind when they reach the termination zone. The speed at which condensin translocates is reduced when it encounters a RNAP. Moving RNAPs can push condensin towards the 3′ of the gene if they represent an obstacle for their translocation (see the Materials and Methods section and Supplemental Data 1). (B) Profile of RNAP mimicking a typical WT-situation of a ∼2 kbp-long gene (see e.g., Figs 1 and 3). (C) Residence time profiles of condensin along the gene when condensin and RNAP move in the same (head-to-tail, full lines) or opposite (head-to-head, dashed lines) direction for two different bypassing rates. (D) Interplay between the translocation of condensin and transcription—backtrack model. (A) This model makes the same basic assumptions as in (A) but RNAP can now dynamically switch between two states: either mobile (mRNAP) or backtracked (bRNAP). The reduction in condensin speed due to collisions with RNAP is stronger with bRNAP than with mRNAP (see the Materials and Methods section and Supplemental Data 1). (E) Rate of backtracking along the gene used in the model. (F, G) Wild-type situation. (F) Density of total RNAP (black line) and bRNAPs (red line) over a ∼2 kbp-long gene. (G) Residence time profiles of condensin along the gene for head-to-tail (full black line) or head-to-head (dashed black line) collisions for a fast bypass rate over mobile RNAP (15 bp/s). The blue full line represents the average between both profiles. (H, I) Over-expression of tfs1DN. (H) Profiles of RNAP (black line) and backtracked RNAPs (red line) obtained by increasing the dwell-time of the backtracked state by 10-fold to mimic the tfs1DN situation. (I) As in (G) but for the tfs1DN-like simulations.
- Figure S3. Mathematical model of the interplay between the translocation of condensin and immobile obstacles bound to chromatin.
(A) Condensin translocates along chromatin from either 5′ to 3′ or 3′ to 5′ at a fast rate (∼1 kbp/s). When it encounters an obstacle bound to chromatin, its translocating speed is strongly reduced (∼10 bp/s). Obstacles can dynamically bind and unbind to chromatin with a typical life-time of 1 min. Simulations of this stochastic system were performed using Gillespie simulations as for the models described in Fig 4A and D (see the Materials and Methods section of the main text). (B) Two toy examples of obstacle occupancy. (C) The residence time of condensin at a given position follows the profile of obstacles occupancy and is symmetric regarding the directionality of condensin.
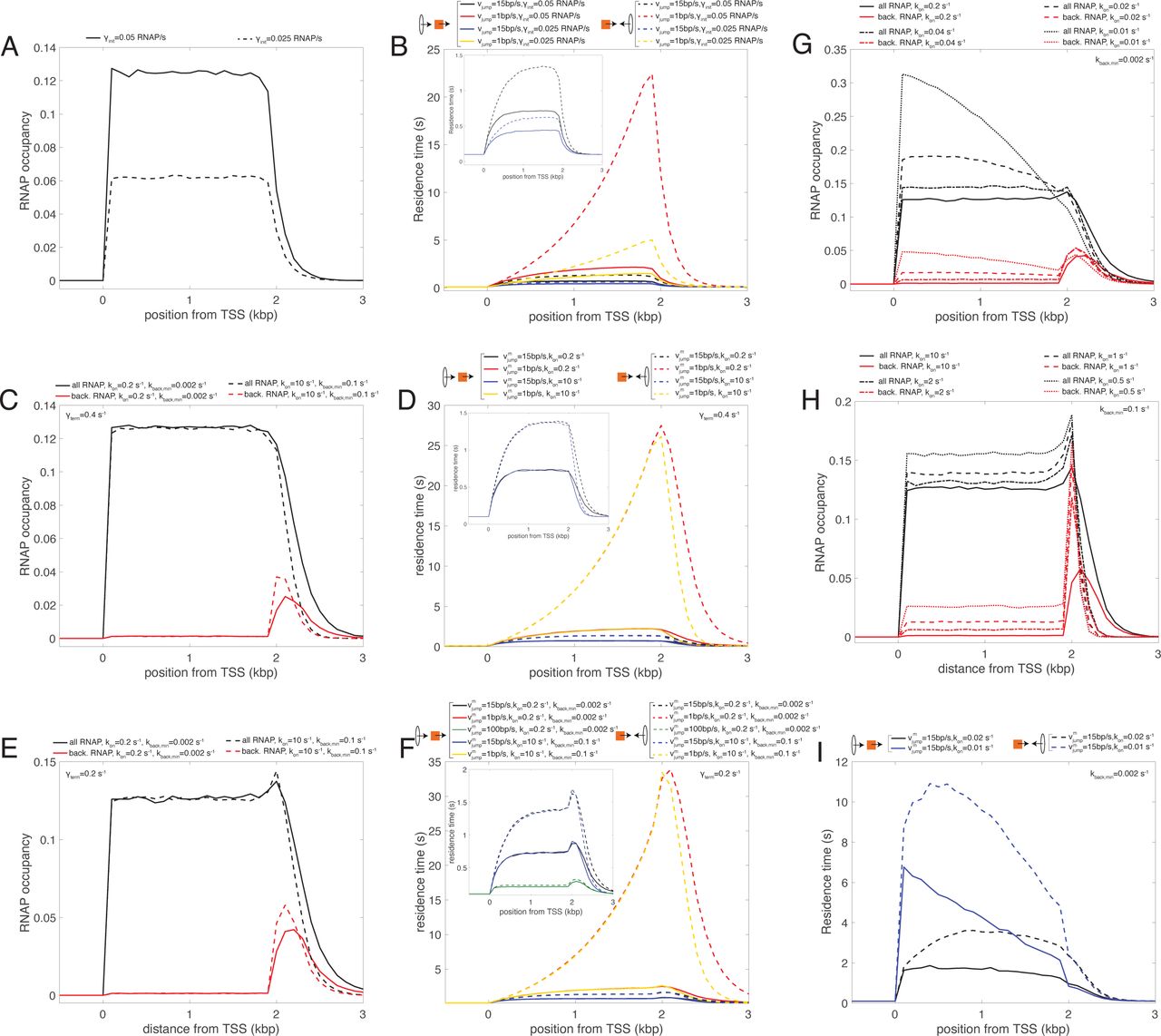
- Figure S4. Analysis of the simple and backtrack models.
(A) RNA polymerase (RNAP) occupancy predicted by the simple model (γterm = vc = 0.4 s−1) for two different values of the initiation rate γinit. (B) Profiles of residence time in the simple model for condensins translocating from 5′ to 3′ (head-to-tail, full lines) or from 3′ to 5′ (head-to-head, dashed lines) for both γinit values shown in (A) (red & black for γinit = 0.05 s−1, blue & yellow for γinit = 0.025 s−1) and for two values of the bypassing rate vjump (black & blue for a “fast” rate, red & yellow for a “slow” rate). The inset magnifies the profiles for the higher vjump value. (C) RNAP occupancy predicted by the backtrack model (γterm = vc = 0.4 s−1, γinit = 0.05 s−1) for “slow” (full lines) or “fast” (dashed lines) switching rates kon and kback,min. Black lines represent the full RNAPs occupancy, whereas the red lines only the bRNAPs. (D) Profiles of residence time for condensins translocating from 5′ to 3′ (head-to-tail, full lines) or from 3′ to 5′ (head-to-head, dashed lines) for parameters corresponding to RNAPs profiles shown in (C) (red & black for “slow” switching rates, blue & yellow for “fast” switching rates) and for two values of the bypassing rate




Supplementary Materials
Supplemental Data 1.
In-depth presentation of the mathematical simulations used in this study.[LSA-2021-01046_Supplemental_Data_1.docx]
Table S2 Primers used in this study.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- Transcription regulates the spatio-temporal dynamics of genes through micro-compartmentalization
- Telomere protein arrays stall DNA loop extrusion by condensin
- Condensin positioning at telomeres by shelterin proteins drives sister-telomere disjunction in anaphase
- RNA Pol II antagonises mitotic chromatin folding and chromosome segregation by condensin
- Condensin positioning at telomeres by shelterin proteins drives sister-telomere disjunction in anaphase
- Context-dependent transcriptional remodeling of TADs during differentiation
- RNA surveillance controls 3D genome structure via stable cohesin-chromosome interaction