




Article Figures & Data
Figures
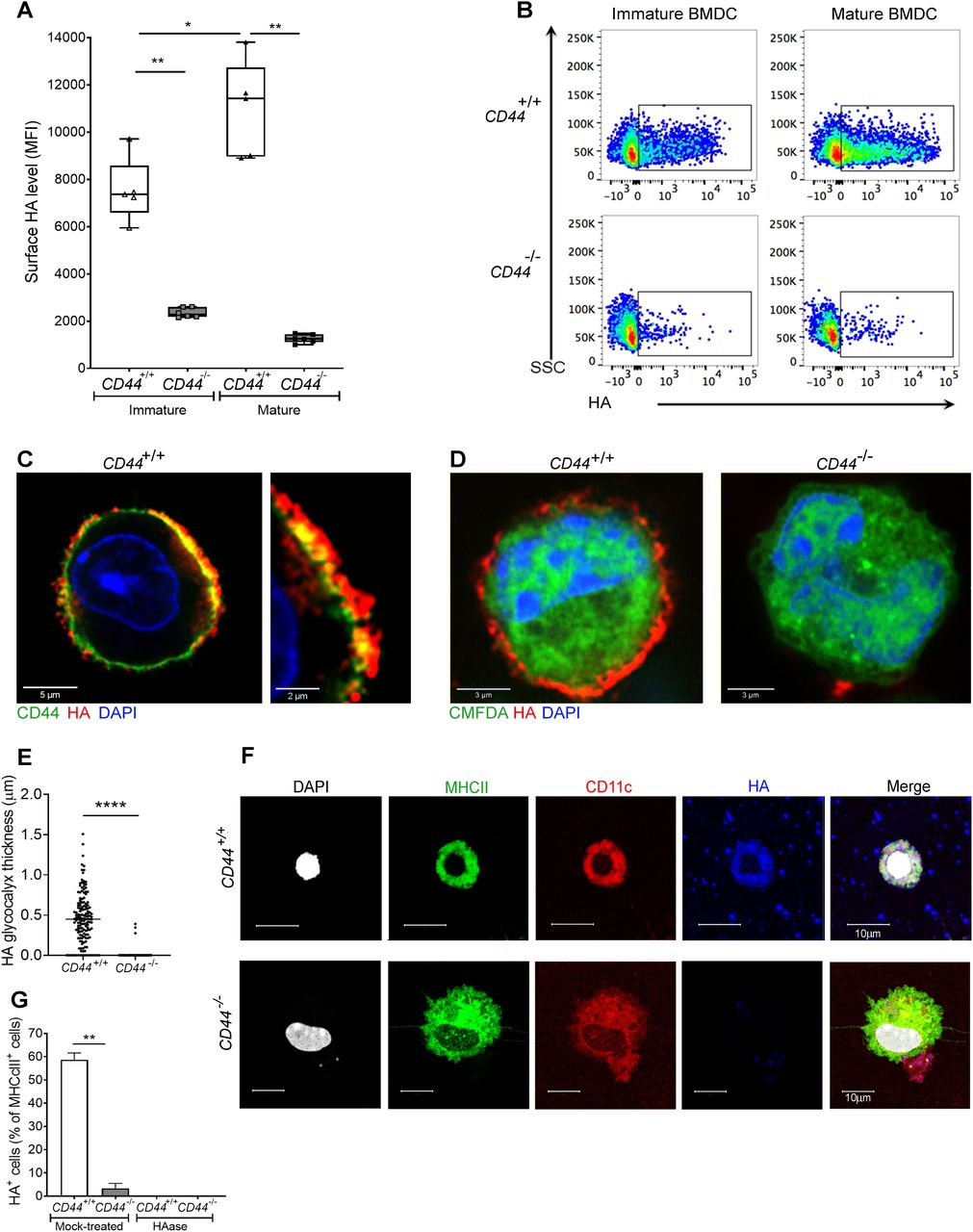
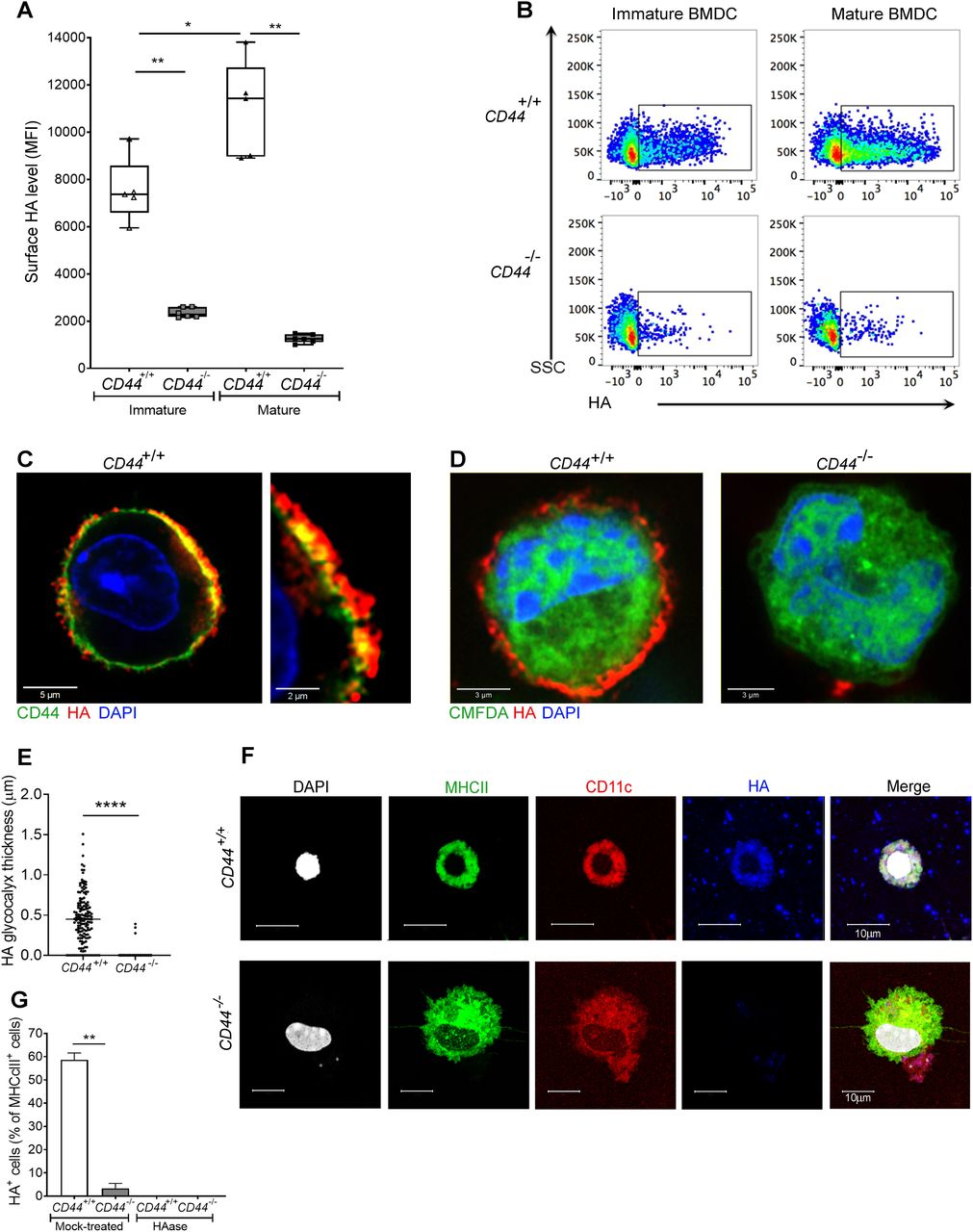
- Figure 1. CD44 anchors the HA glycocalyx to the surface of both immature and mature DCs.
(A) HA on intact BMDCs prepared from CD44+/+ and CD44−/− littermates, as detected using bVG1 with streptavidin-AF647 by flow cytometry. Data in box plots represent the median (center bar) ± s.e.m. (whiskers), n = 5, 6 mice. Data are one representative experiment from three. (B) Representative dot plots of bVG1-streptavidin-AF647, to indicate levels of cell surface HA. (C) Visualization of the HA glycocalyx by bVG1–streptavidin–AF647 (red) on an individual PFA-fixed CD44+/+ BMDC, with CD44 detected by mAb IM7 (green) and nucleus counterstained with DAPI (blue), as observed by confocal microscopy with Airyscan detection, with digital zoom (right panel) to show dimensions of the extended pericellular coat. (D) Confocal microscopy of fixed CD44+/+ and CD44−/− BMDCs labeled with CMFDA green tracker dye, bVG1-streptavidin-AF647 (red), and counterstained with DAPI (blue). (E) Thickness of the HA glycocalyx on fixed CD44+/+ and CD44−/− BMDCs, as measured from Airyscan images using Image J (n = 5, 5 cells, data combined from three experiments). (F) Detection of HA (blue) on migratory MHC class II+ (green) CD11c+ (red) dermal DCs, with nuclei counterstained with DAPI (gray), following egress from tissue during ex vivo culture. (G) Percentage of MHC class II+ dermal DCs scored as surface HA+ by microscopy (n = 6, 6 mice, data combined from two experiments), following fixation of freshly resected ear skin in PFA and incubation for 2 h at 37 °C ± hyaluronidase (HAase), before immunostaining *P < 0.05, **P < 0.01, ****P < 0.0001, Mann–Whitney U-test.
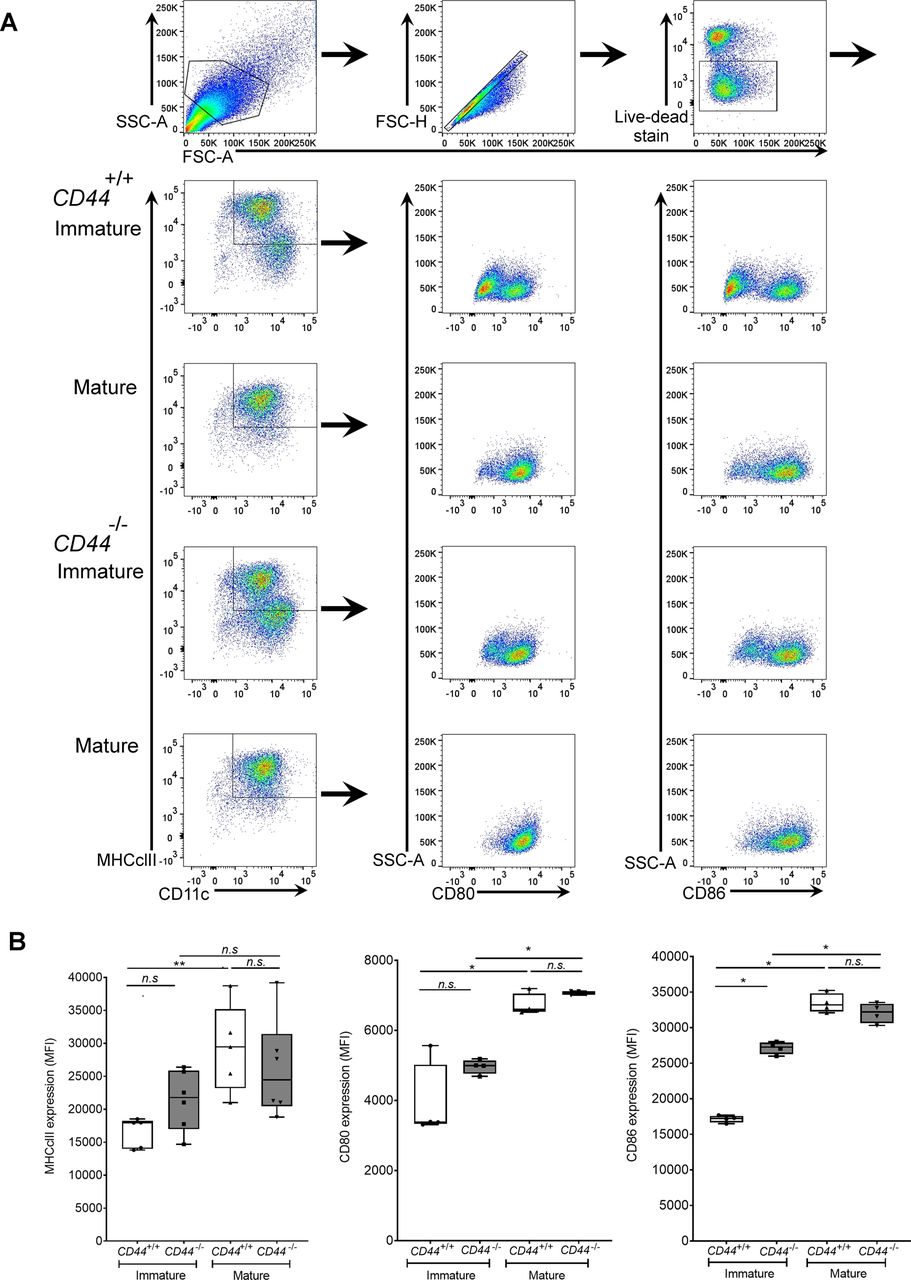
- Figure S1. CD44 deficiency does not affect expression of DC activation markers.
BMDCs were prepared from CD44+/+ and CD44−/− littermates and stimulated for 24 h with LPS to induce maturation. Surface expression of MHC class II, CD80, and CD86 was measured by flow cytometry. (A) Flow cytometry gating and representative dot plots. (B) Data in box plots represent the mean (center bar) ± s.e.m. (whiskers) (n = 5, 6 mice).
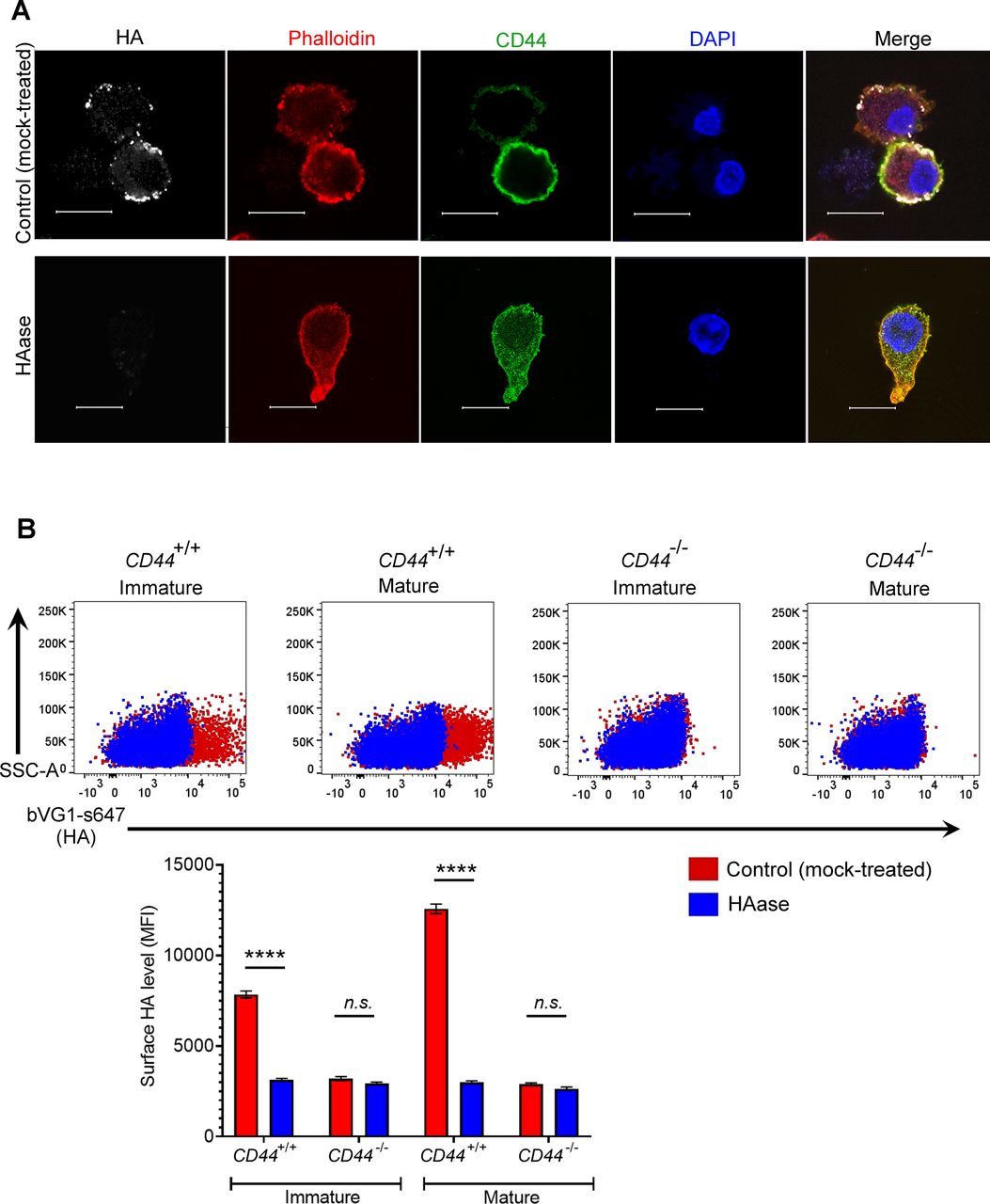
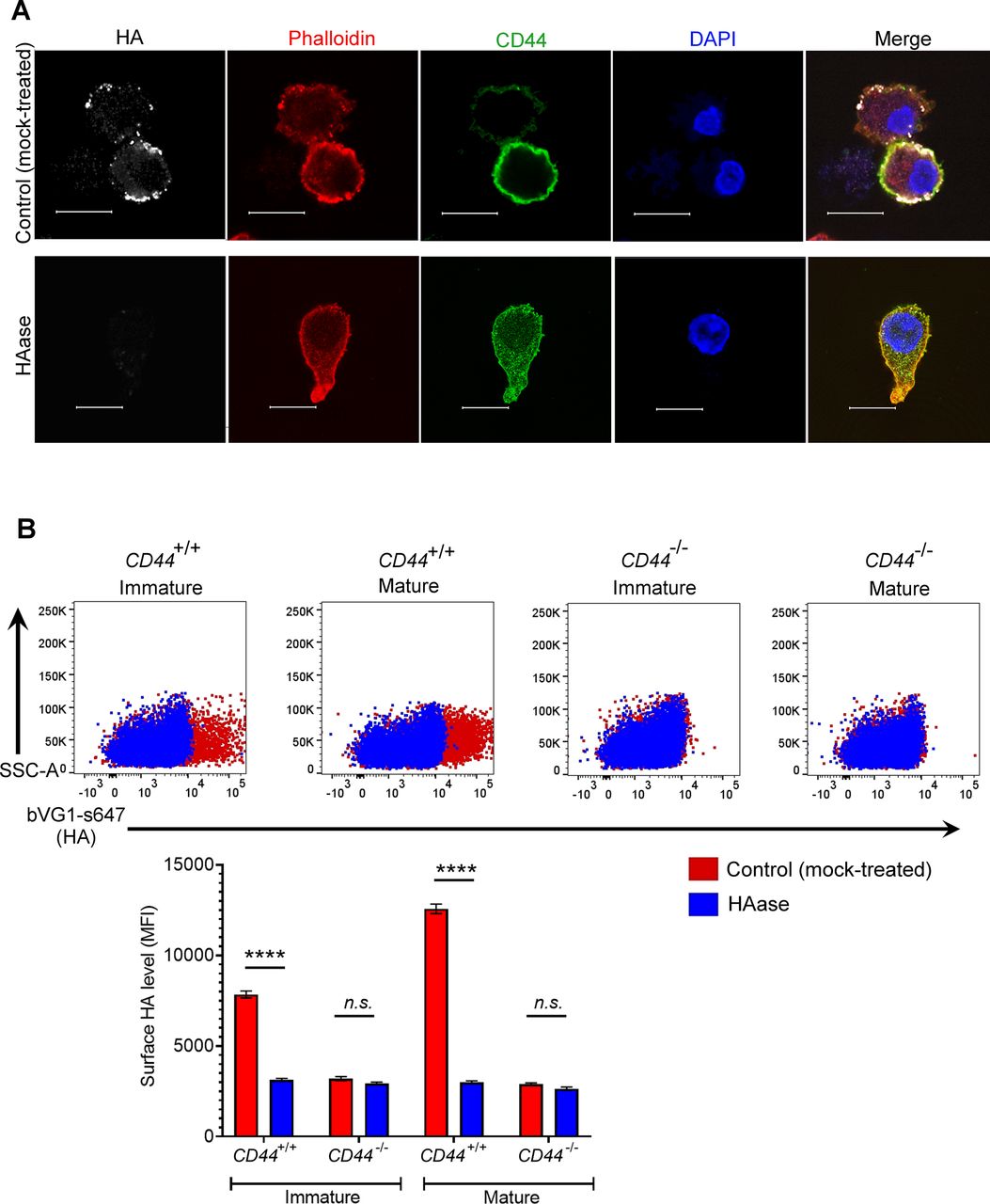
- Figure S2. Binding specificity of the HA probe bVG1.
(A) Mock- or hyaluronidase (HAase)-treated mature wild-type BMDCs were permeabilized then incubated with bVG1 and streptavidin-AF647, immunostained with anti-CD44 (green) and counterstained with phalloidin (red) and DAPI (blue) before Airyscan confocal imaging. Scale bar = 10 μm. (B) Surface HA was detected on intact mock- or hyaluronidase (HAase)-treated cells by flow cytometry, using bVG1 and streptavidin-AF647. Representative dot plots are shown, with data in box plots showing the mean fluorescence intensity ± s.e.m. (n = 4).
- Figure S3. Reduced HA levels in the dermis of CD44-deficient mice.
(A) Detection of HA (blue) on MHC class II+ dermal DCs (green), indicated by arrow, in skin excised from CD44+/+ and CD44−/− mice, as shown by whole-mount immunostaining and confocal microscopy, with lymphatic vessels labeled with anti–LYVE-1 (red) and HA by bVG1/streptavidin–AF647 (blue), scale bar = 20 μm. (B, C) Measurement of total levels of pericellular and extracellular HA in dermis of CD44+/+ and CD44−/− mice, as shown by whole-mount immunostaining and confocal microscopy, with lymphatic vessels labeled with anti–LYVE-1 (red) and HA by bVG1/streptavidin-AF647 (blue), scale bar = 100 μm (B). (C) Quantitation of HA levels and LYVE-1 expression (C) using Image J software. *P < 0.05, Mann–Whitney U-test. Data are the mean (center bar) ± s.e.m. (whiskers) (n = 4 mice). Data are representative of one experiment from three separate experiments.
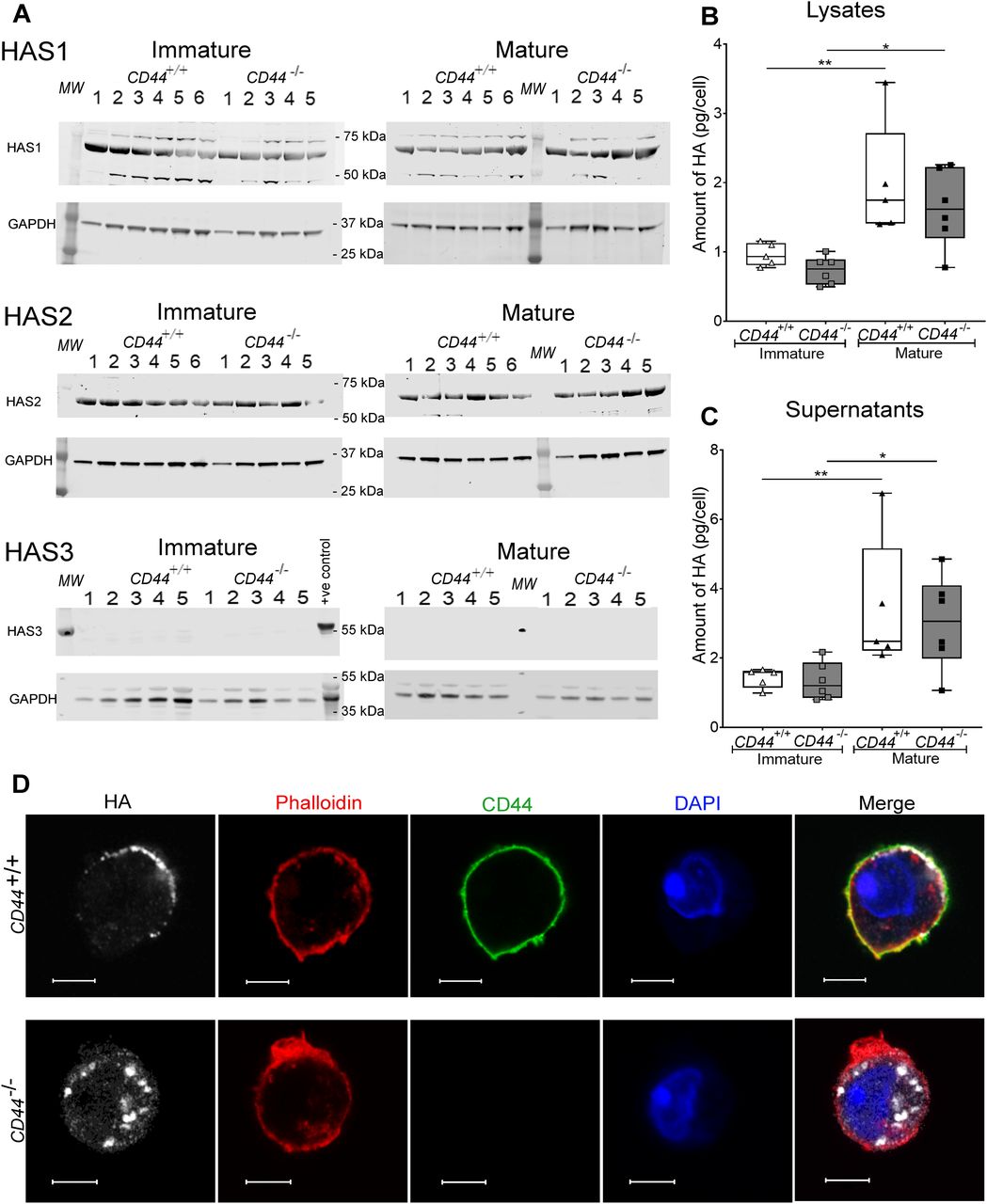
- Figure 2. CD44−/− BMDCs exhibit no defect in HA synthesis.
(A) Cell lysates of immature and mature BMDCs from either five or six individual CD44+/+ and five CD44−/− littermates were resolved by SDS–PAGE and probed by Western blotting to detect HAS enzymes HAS1-3 and GAPDH (as a loading control). Representative blots are shown, from one of two separate experiments. Lysate from wild-type mouse skin was included as a positive control for HAS3. (B, C) Quantitation of HA in lysates (B) and supernatants (C) of immature and LPS-matured BMDCs, as determined by ELISA. Data represent the median (center bar) ± s.e.m. (whiskers), n = 5 for CD44+/+ and 6 for CD44−/− mice, one representative experiment from three separate experiments. (D) Detection of HA in permeabilized BMDCs prepared from CD44+/+ and CD44−/− littermates, using bVG1 and streptavidin–AF647 (gray), immunostaining for CD44 (green), and counterstaining with phalloidin (red) and DAPI (blue). Scale bars = 5 μm.
- Figure S4. CD44 deficiency does not affect expression of HAS enzymes.
HA synthase enzymes HAS1-3 and GAPDH in BMDC lysates were resolved by SDS–PAGE and probed using Western blotting. Representative blots of BMDCs sampled from six individual CD44+/+ and five CD44−/− littermates (shown in Fig 2). Expression of HAS1 and HAS2 relative to GAPDH was calculated for each blot. Data are representative of one experiment from three separate experiments.
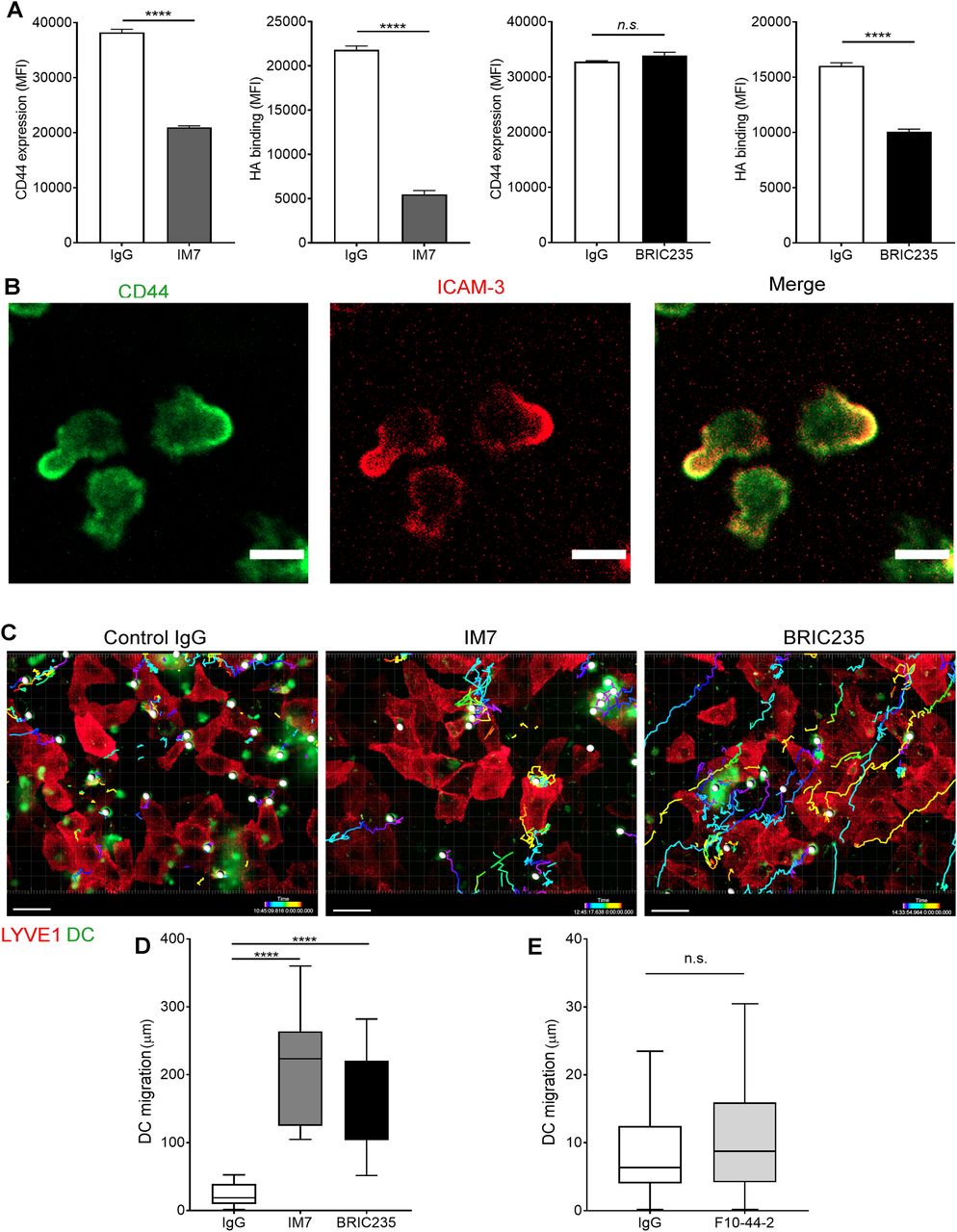
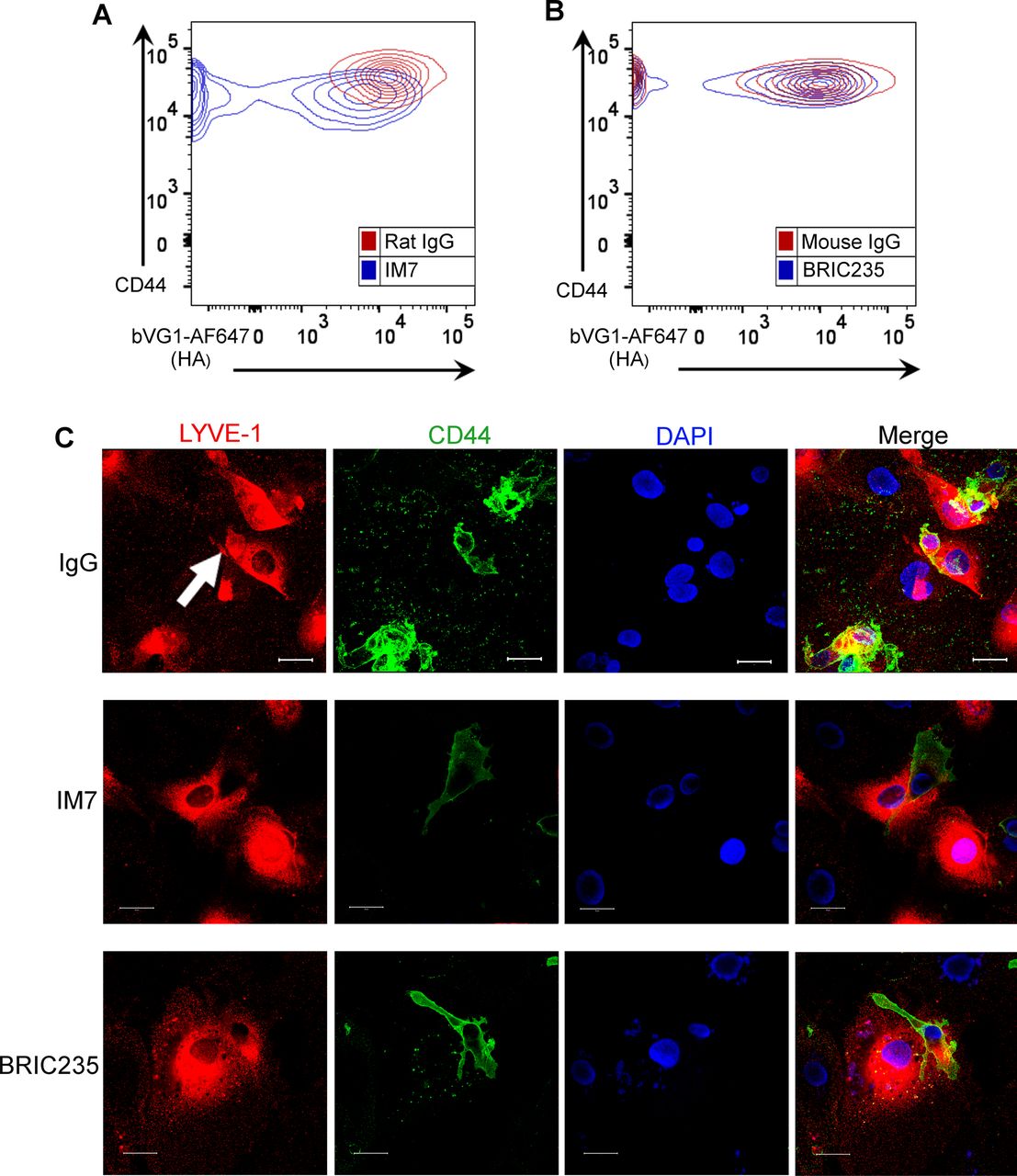
- Figure 3. Disrupting CD44-HA binding impairs DC interactions with human lymphatic endothelial cells (hLECs).
(A) Surface levels of CD44 and HA on LPS-matured human monocyte-derived DC (MDDC) after incubation with control IgG, or anti-CD44 mAbs IM7 and BRIC235, as measured by flow cytometry. ****P < 0.0001, unpaired t test. Data are the mean ± s.e.m. (n = 3), one representative experiment of three. (B) Single time point from a video of MDDCs labeled with FITC-conjugated anti-CD44 (mAb F10.44.2, green) and APC-conjugated anti–ICAM3 (red) crawling on unstained hLEC monolayers. Scale bar = 10 μm. (C, D, E) Migration of mAb-treated MDDCs on monolayers of hLECs, imaged by spinning disc confocal microscopy over 4 h, with dragon tail tracks color-coded to indicate time progression (C) and quantitated using Imaris software (D, E). Scale bar = 20 μm, ****P < 0.0001, Mann–Whitney U-test. Data are the median (center bar) ± s.e.m. (whiskers), data combined from three experiments.
- Figure S5. Disrupting CD44–HA interactions on monocyte-derived DCs (MDDCs) with mAbs prevents formation of lymphatic endothelial transmigratory cups.
(A, B) Representative contour plots from flow cytometry data shown in Fig 3A, showing surface levels of CD44 and HA on LPS-matured human MDDC after incubation with control IgG, or anti-CD44 mAbs IM7 (A) or BRIC235 (B). (C) Confocal microscopy of human lymphatic endothelial cell monolayers co-cultured with mature MDDC for 3 h in the presence of either IgG, IM7, or BRIC235, then immunostained with anti–LYVE-1 (red) and anti-CD44 (green), with nuclei counterstained using DAPI (blue). Scale bar = 20 μm.
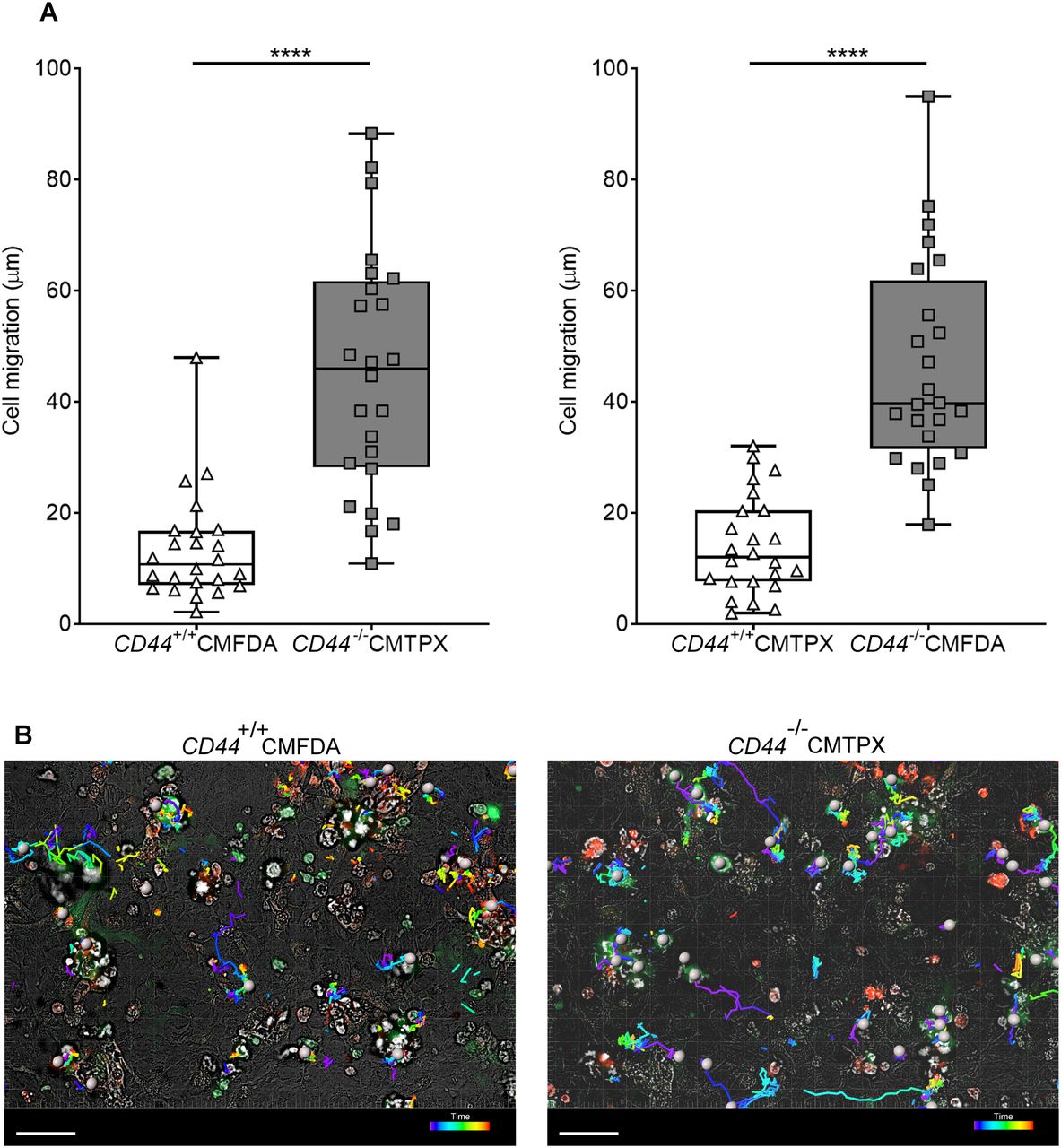
- Figure 4. CD44 deficiency impairs BMDC interactions with mouse lymphatic endothelial cells.
Tracking migration of LPS-matured BMDCs from CD44+/+ and CD44−/− mice, labeled with CMFDA green or CMTPX red tracker dyes, applied to monolayers of mouse lymphatic endothelial cells and imaged by spinning disc confocal microscopy over 4 h. (A) Lengths of migratory paths were tracked using Imaris software. ****P < 0.0001, Mann–Whitney U-test. Data are the median (center bar) ± s.e.m. (whiskers) eight cells/field of view, three fields of view, data combined from three experiments. (B) Representative images of dragon tail paths to indicate time are shown for CD44+/+ BMDCs (CMFDA, green) and CD44−/− BMDCs (CMPXT, red) with bright-field, and cells indicated by gray spheres. Scale bar = 50 μm.
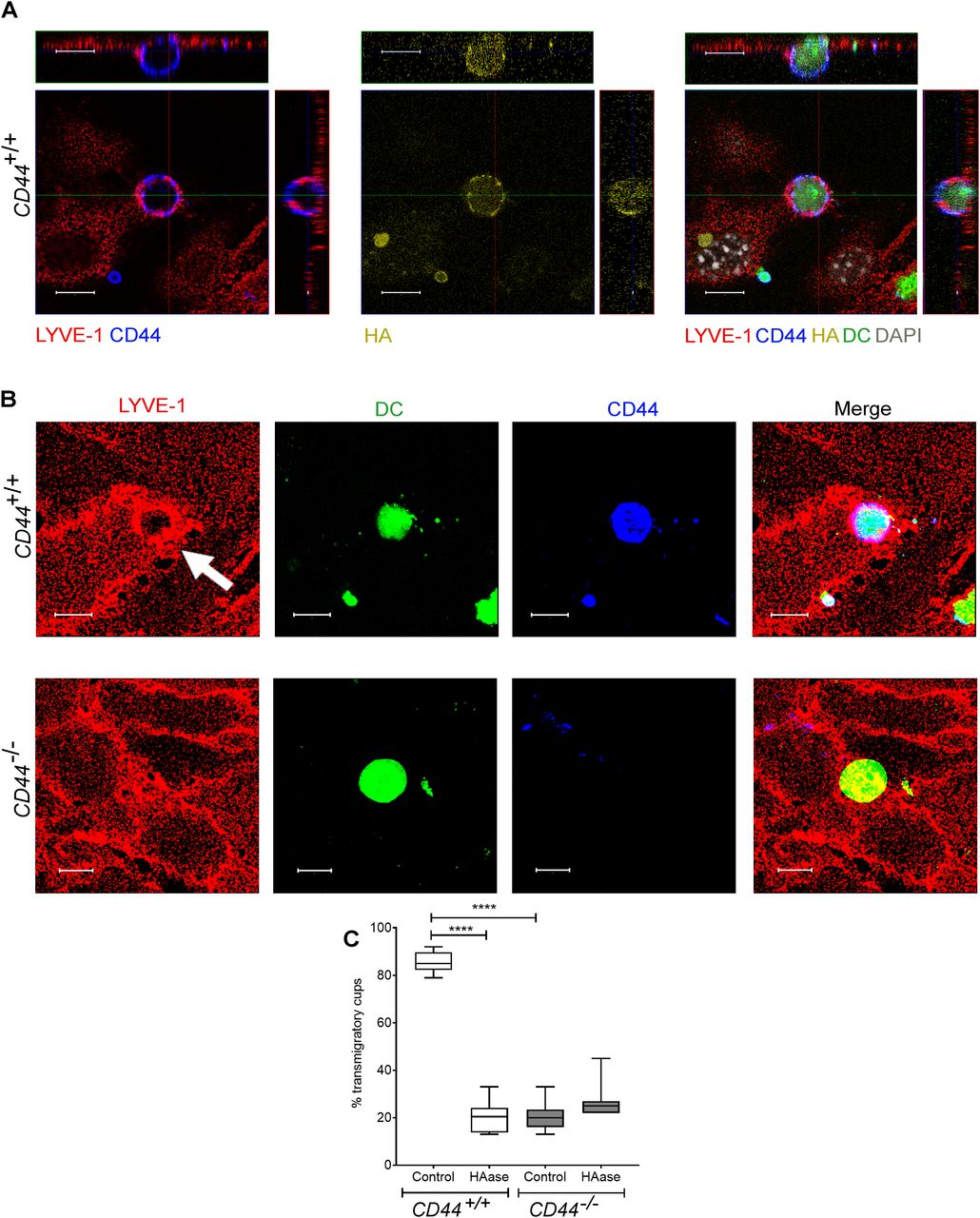
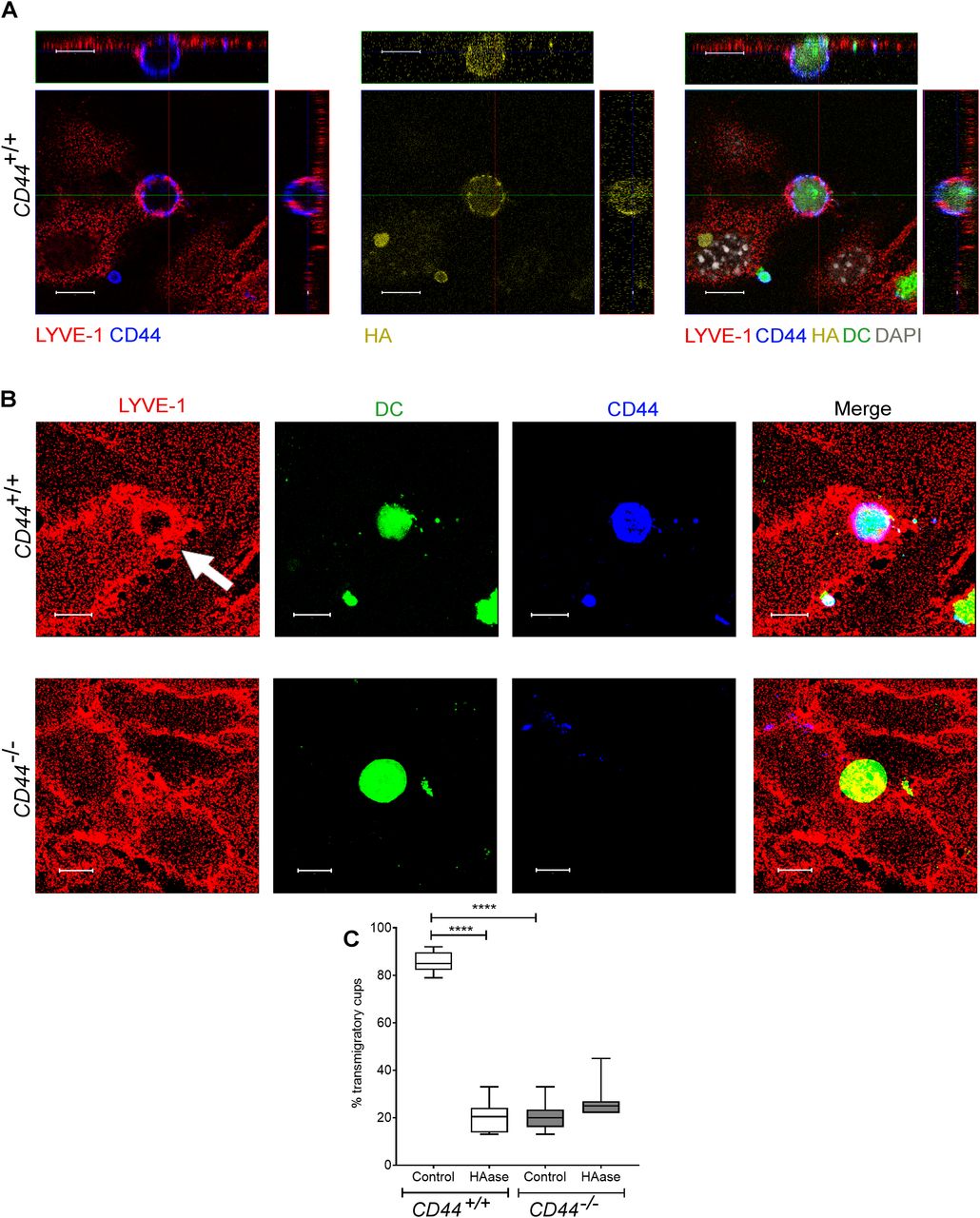
- Figure 5. CD44 is required for efficient formation of lymphatic endothelial transmigratory cups.
(A) Confocal microscopy orthogonal view images of cultured primary mouse lymphatic endothelial cell (mLEC) monolayers immunostained with anti–LYVE-1 (red) viewed 3 h after the addition of LPS-matured, fluorescently labeled (green) CD44+/+ BMDC immunostained with anti-CD44 (blue), and bVG1 (yellow) and counterstained with DAPI (gray), scale bar = 10 μm. (B) Three-dimensional rendering of confocal images of cultured primary mLEC monolayer immunostained with anti–LYVE-1 (red) viewed 3 h after the addition of fluorescently labeled (green) CD44+/+ or CD44−/− BMDC immunostained with anti-CD44 (blue). An individual LYVE-1-lined transmigratory cup is indicated by arrow. Scale bar = 10 μm. (C) Quantitation of microscopy images to show the number of LYVE-1+ transmigratory cups associating with adherent DCs, after 2-h preincubation with HAase and 3-h co-culture of mLEC monolayers with either mature CD44+/+ or CD44−/− BMDC. ****P < 0.0001, Mann–Whitney U-test. Data are the median (center bar) ± s.e.m. (whiskers) (n = 10 fields of view), data combined from three experiments.
- Figure 6. Impaired trafficking of endogenous dermal DCs to draining LNs in CD44-deficient mice.
(A, B) Recovery of endogenous DCs from draining inguinal and axillary LNs, 24 h after topical application of oxazolone and FITC, as measured by flow cytometry. (A, B) Live CD45+ CD11c+ FITC+ cells (A) were further gated according to expression of MHC class II and CD103 (B). (C) Total cellularity of LNs was measured by counting cells with a hemocytometer. *P < 0.05, Mann–Whitney U-test. Data represent the mean (center bar) ± s.e.m. (whiskers) (n = 5 mice), one representative experiment of three. (D) Representative contour and dot plots, showing percentage of cells in each gate.
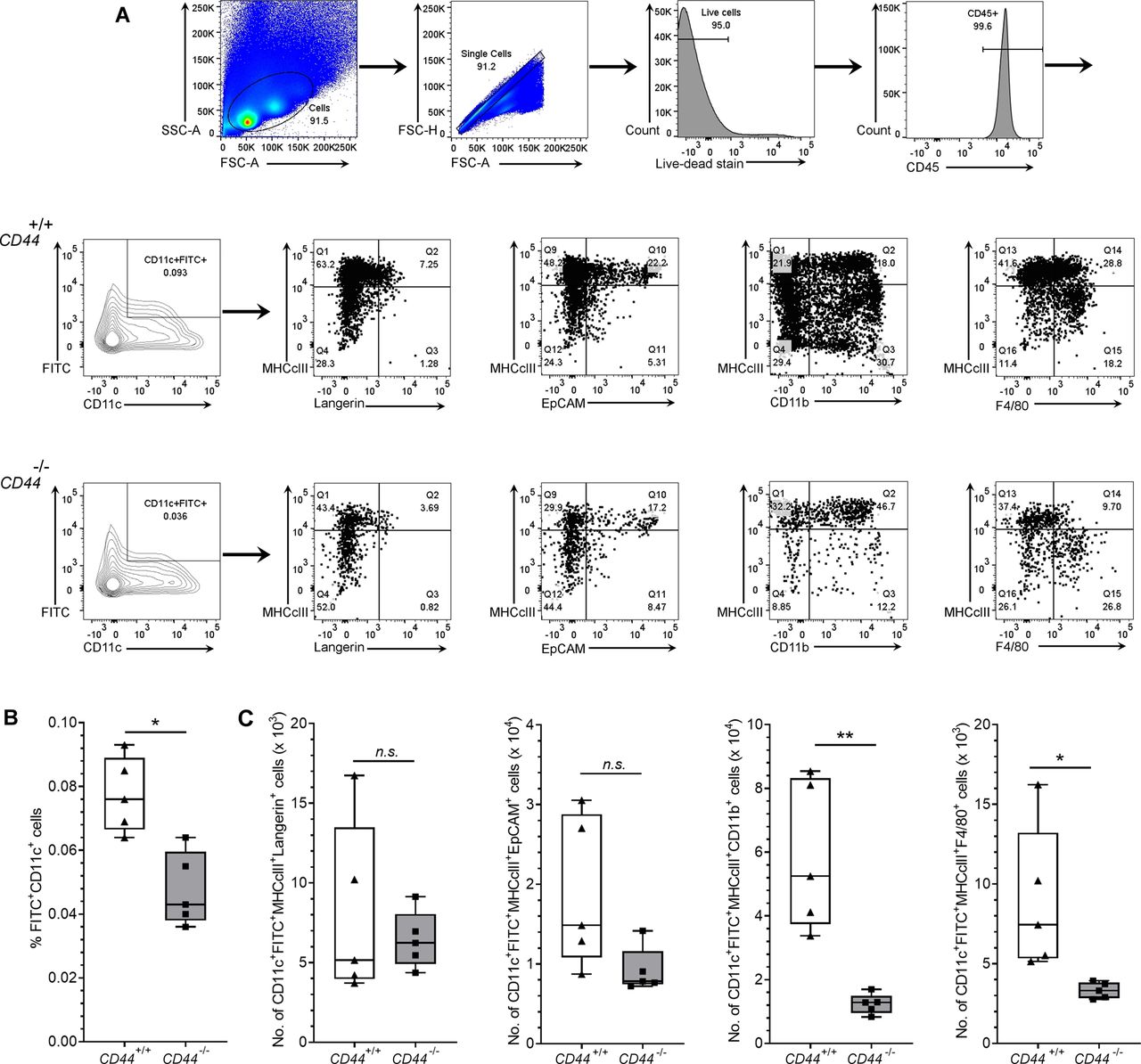
- Figure S6. Impaired trafficking of dermal DCs to draining LNs in CD44-deficient mice.
(A, B, C) Recovery of endogenous DCs from draining inguinal and axillary LNs, 24 h after topical application of oxazolone and FITC, as measured by flow cytometry. (A) Gating strategy for data in Fig 6 is shown, where live CD45+ CD11c+ FITC+ cells were further gated according to expression of MHC class II and either Langerin, EpCAM, CD11b or F4/80, with representative contour and dot plots, showing percentage of cells in each gate (A). (B, C) Bar and whiskers plots showing percentage of LN cells that are FITC+ CD11c+ (B) and absolute numbers of CD11c+ MHC class II+ FITC+ cells in LN expressing subset markers Langerin, EpCAM, CD11b, and F4/80 (C). *P < 0.05, **P < 0.01, Mann–Whitney U-test. Data represent the mean (center bar) ± s.e.m. (whiskers) (n = 5 mice), one representative experiment of three.
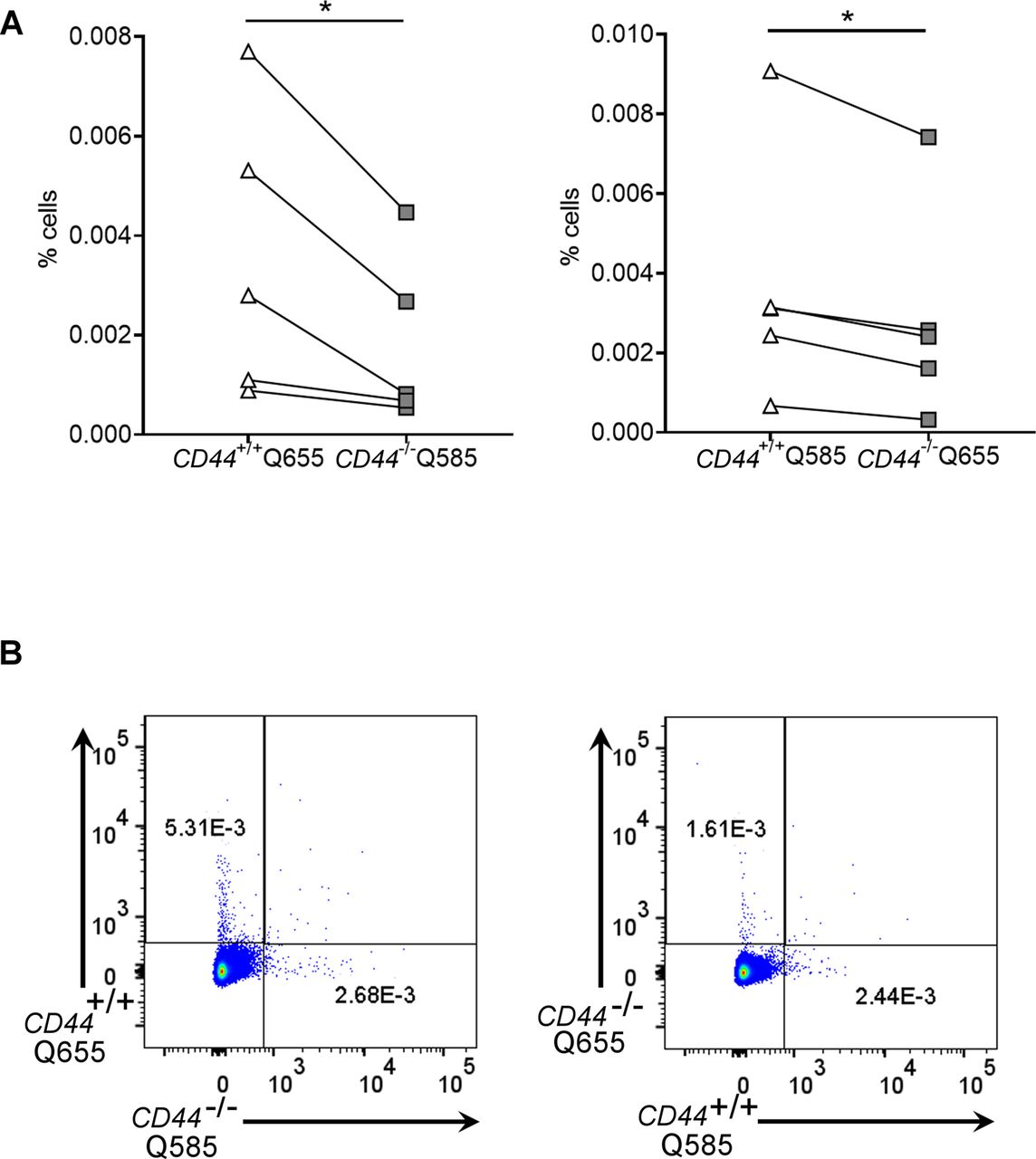
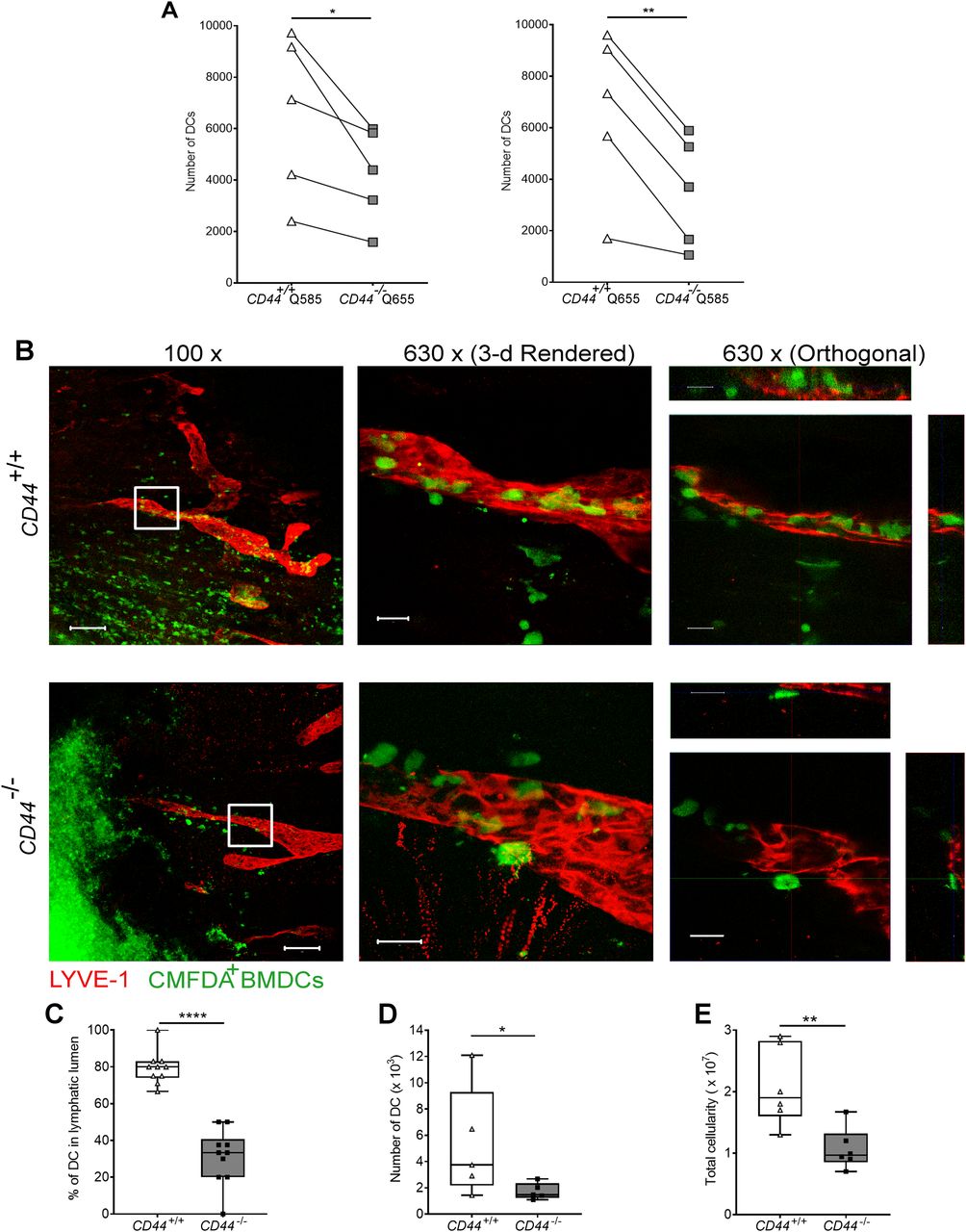
- Figure 7. Impaired entry and trafficking of CD44−/− BMDCs in dermal lymphatic vessels.
(A) Comparison of lymphatic trafficking of LPS-matured CD44+/+ and CD44−/− BMDCs, differentially labeled with Q-dot 585 and 655, respectively, or vice versa, 24 h after intradermal co-injection into oxazolone-painted skin of CD44+/+ mice. Recovery of BMDCs in draining cervical LNs was measured by flow cytometry, *P < 0.05, **P < 0.01, Paired t tests. (B) Entry of CMFDA-labeled CD44+/+ and CD44−/− BMDCs into dermal afferent lymphatics immunostained with anti–LYVE-1, following topical administration of oxazolone. Panels show 3-D rendering of z-stacks at low magnification (left; 100×, scale bars = 100 μm), and higher magnification (middle; 630×, scale bars = 20 μm) with orthogonal sections (right). (C) Numbers of BMDCs inside lymphatic vessel lumens, expressed as a percentage of the number of lymphatic vessel-associated BMDCs. Data combined from three experiments, ****P < 0.0001, Mann–Whitney U-test. (D, E) Recovery of intradermally injected CMFDA-labeled BMDCs (D) and overall cellularity (E) in draining cervical LNs 24 h after topical application of oxazolone and adoptive transfer of BMDCs, as measured by flow cytometry. *P < 0.05, **P < 0.01, Mann–Whitney U-test. Data are the median (center bar) ± s.e.m. (whiskers) (n = 5 mice). One representative experiment of three.
- Figure S7. Impaired lymphatic trafficking of CD44−/− BMDCs from dermis.
(A, B) Comparison of lymphatic trafficking of LPS-matured CD44+/+ and CD44−/− BMDCs, differentially labeled with Qdot 585 and 655, respectively, or vice versa, 24 h after intradermal co-injection into oxazolone-painted skin of CD44+/+ mice. (A, B) Recovery of BMDCs in draining cervical LNs was measured by flow cytometry and expressed as a percentage of total number of cells (A), with representative dot plots shown from data in Fig 7 (B). *P < 0.05, Paired t tests.
- Figure 8. DC adhesion and transendothelial migration are dependent on CD44 and HA.
(A, B, C) Adhesion of LPS-matured CMFDA-labeled CD44+/+ and CD44−/− BMDCs to mouse lymphatic endothelial cell (mLEC) monolayers after 3 h incubation, as assessed by fluorescence plate reader, comparing (A) immature and mature BMDC, (B), mature BMDC after 2-h incubation with or without hyaluronidase (HAase) and (C) mature wild-type BMDC in the presence of exogenously applied high molecular weight HA. (D) Immunostaining and confocal microscopy of mLEC monolayers following 3 h co-culture with CD44+/+ or CD44−/− LPS-matured CMFDA-labeled BMDC (green), with anti–LYVE-1 (red) and nuclei counterstained with DAPI (blue). Scale bar = 100 μm. (E, F, G) Transmigration of LPS-matured CMFDA-labeled CD44+/+ and CD44−/− BMDCs through either mLEC monolayers cultured on the undersurface of Transwell filters in the presence of control rat IgG or anti–LYVE-1 neutralizing mAb 2125 where indicated (E, F) or across fibroblast monolayers cultured on the upper surface of filters (G) measured over a 5-h period by fluorescence plate reader. *P < 0.05, Mann–Whitney U-test. Data are the mean (center bar) ± s.e.m. (whiskers) (n = 4), representative experiments of three separate experiments.
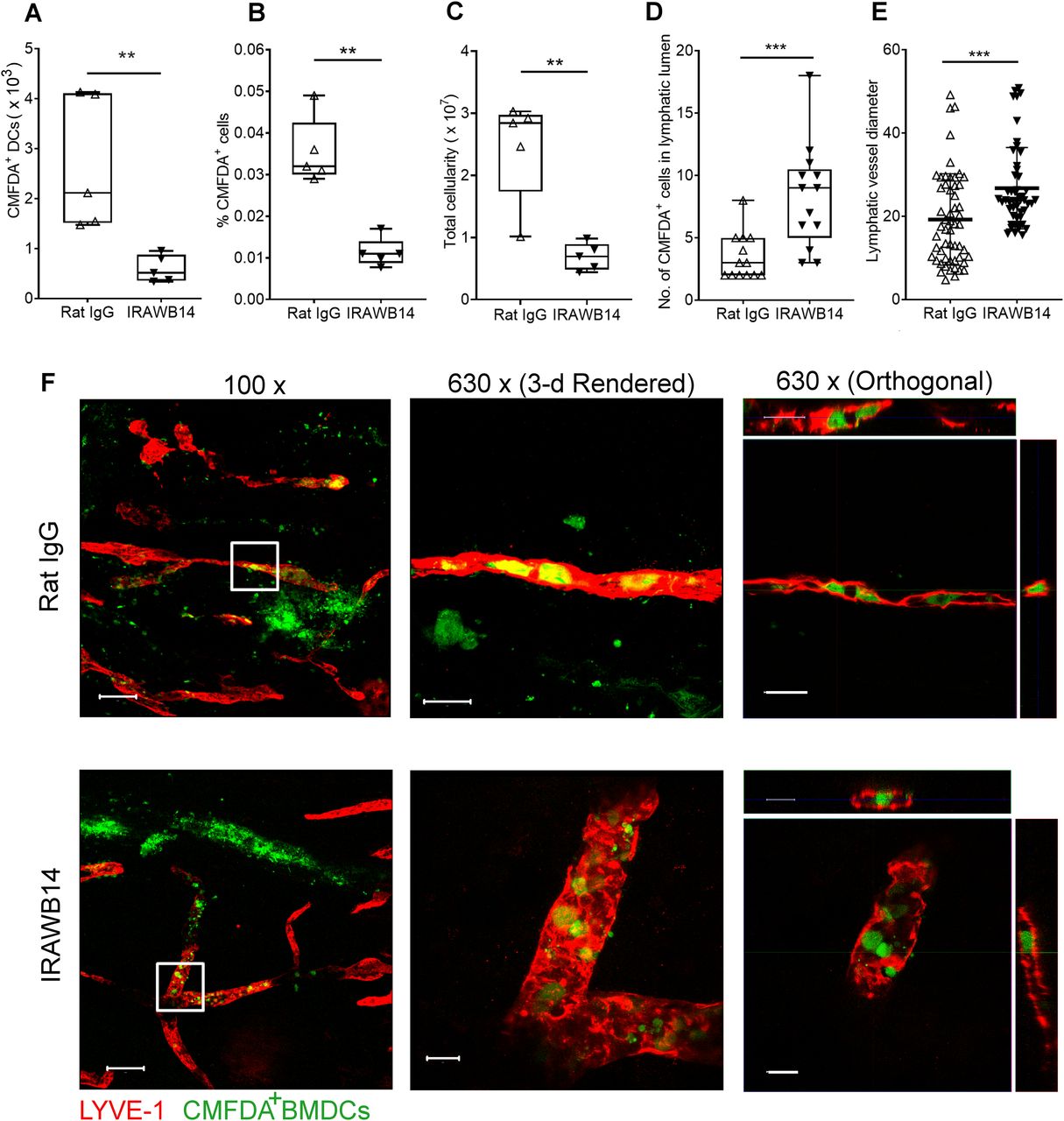
- Figure 9. mAb-induced enhancement of CD44-HA binding increases BMDC adhesion to mouse lymphatic endothelial cell (mLEC) and impairs transmigration.
(A, B) Capacity of LPS-matured CD44+/+ and CD44−/− BMDCs to bind endogenously synthesized HA (A) or exogenously added HA (B), assessed by incubation with bVG1 and streptavidin–AF647 or bHA and streptavidin–AF647, respectively, with quantitation by flow cytometry and showing representative contour plots. (C, D) Adhesion of LPS-matured CMFDA-labeled wild-type BMDCs (green) to mLEC monolayers after 3-h incubation in the presence of either rat IgG or the CD44 potentiating antibody IRAWB14, as assessed by fluorescence plate reader (C) and confocal microscopy (D), with mLEC immunostained for LYVE-1 (red) and counterstained with DAPI (blue). Scale bar = 50 μm. (E, F) Effect of IRAWB14 or control rat IgG on transmigratory cup formation in mLEC monolayers co-cultured for 3 h with LPS-matured CMFDA-labeled BMDCs (green) and immunostained for LYVE-1 (red), with bound IRAWB14 (detected with anti-rat IgG, blue), shown in (E), scale bar = 20 μm, and the percentage of adherent BMDCs associated with LYVE-1+ transmigratory cups in each condition shown in (F). (G) Effect of IRAWB14 mAb or control rat IgG on basolateral-to-luminal transmigration of LPS-matured CMFDA-labeled wild-type BMDCs across mLEC monolayers grown on the undersurface of Transwell filters, measured over a 5-h period by fluorescence plate reader. *P < 0.05, Mann–Whitney U-test. Data are the mean (center bar) ± s.e.m. (whiskers) (n = 4), one representative experiment of three.
Source data are available for this figure.
- Figure S8. Effects of mAb IRAWB14 on BMDC viability and migration.
(A) Flow cytometry to assess mAb binding and viability of LPS-matured wild-type BMDCs after 24-h in vitro culture with either control rat IgG or IRAWB14, showing gating strategy, viability according to dye exclusion, and recognition of surface bound mAb using anti-rat IgG fluorescently conjugated to Alexa Fluor 488. (B) Representative flow cytometry dot plots of data in Fig 10A and B, to show recovery of CMFDA+ BMDCs from draining cervical LNs 24 h after intradermal injection into oxazolone-painted skin. Numbers in dot plots indicate the percentage of cells in the gate.
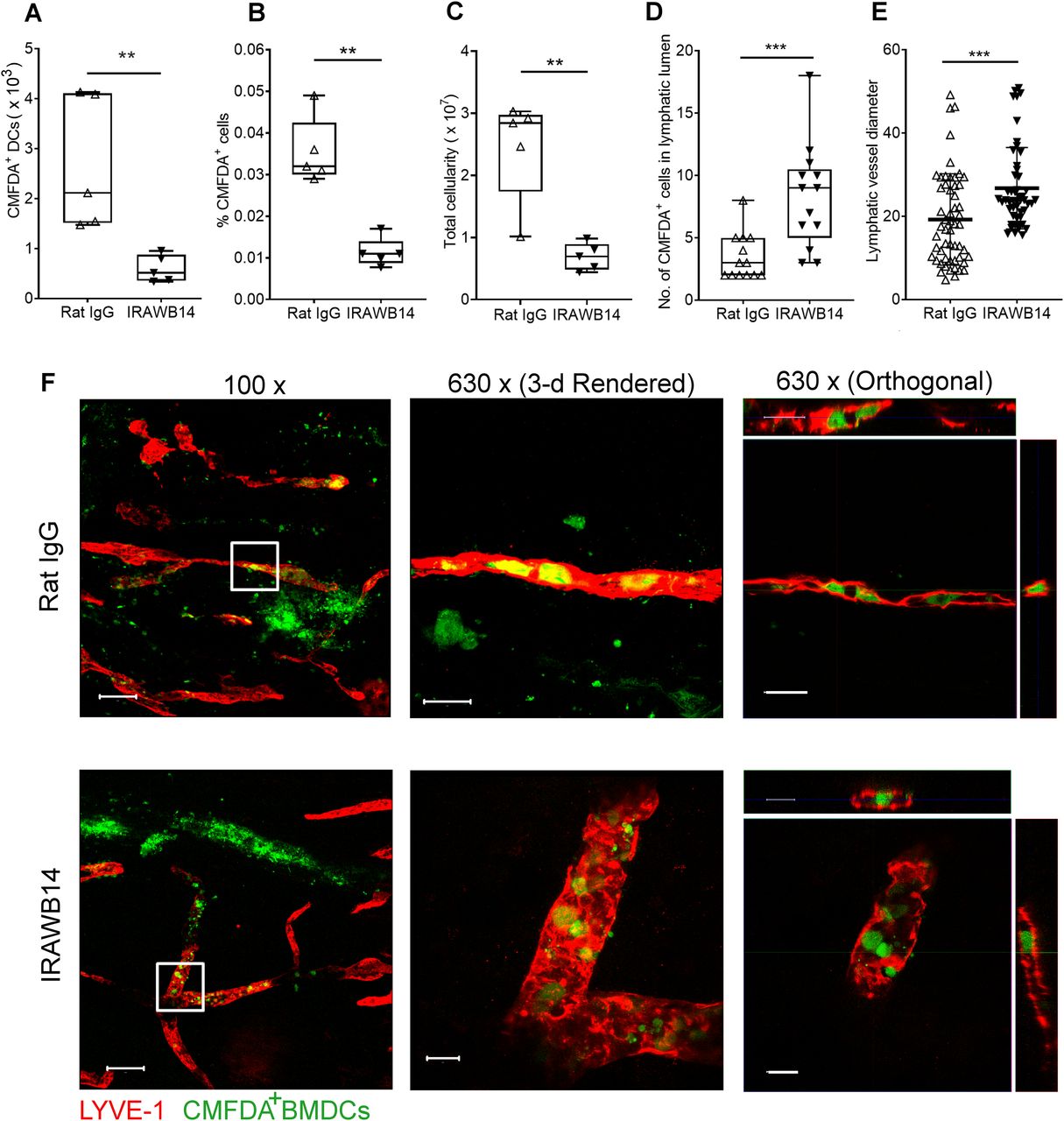
- Figure 10. Impaired lymphatic trafficking of DCs after mAb-induced enhancement of CD44-HA binding.
(A, B, C) Effect of IRAWB14 or control rat IgG on recovery of LPS-matured CMFDA-labeled wild-type BMDCs from draining cervical LNs 24 h after intradermal injection into oxazolone-painted skin, as measured by flow cytometry and expressed as (A) numbers of CMFDA+ BMDCs, (B) percentage of CMFDA+ cells and (C) total LN cellularity. (D) Numbers of BMDCs detected inside the lumen of dermal lymphatic vessels, expressed as a percentage of the total number of lymphatic vessel-associated BMDCs. (E) Lymphatic vessel diameter 24 h after adoptive transfer of IgG- and IRAWB14-treated BMDCs, five measurements/vessel, five vessels/mouse. (F) Entry of CMFDA-labeled IgG- and IRAWB14-treated wild-type BMDCs (green) into dermal afferent lymphatic capillaries immunostained with anti–LYVE-1 (red), following oxazolone skin painting. Panels show 3-D rendering of z-stacks at low magnification (left; 100×, scale bars = 100 μm), and higher magnification (center; 630×, scale bars = 20 μm) with orthogonal sections (right). **P < 0.01, ***P < 0.001, Mann–Whitney U-test. Data are the mean (center bar) ± s.e.m. (whiskers) (n = 5 mice), one representative experiment of three.








Supplementary Materials
- Video 1
Visualization of monocyte-derived DCs crawling and adhering to human lymphatic endothelial cell monolayer. Representative spinning disc confocal microscopy time-lapse of monocyte-derived DC labeled with CMFDA green tracker dye, crawling on a monolayer of confluent human lymphatic endothelial cell labeled with anti–LYVE-1 (mAb6A) conjugated with DyLight 650 fluorophore (red), in the presence of control IgG. Scale bar = 100 μm, time (mins:sec), 20 fps. Video corresponds to the images in Fig 3C and D.Download video
- Video 2
LYVE-1 rearrangements around an adherent monocyte-derived DC on human lymphatic endothelial cell monolayer. Representative spinning disc confocal microscopy time-lapse of monocyte-derived DC labeled with CMFDA green tracker dye, crawling on a monolayer of confluent human lymphatic endothelial cell labeled with anti–LYVE-1 (mAb 6A) conjugated with DyLight 650 fluorophore (red), in the presence of control IgG. Scale bar = 20 μm, time (mins:sec), 20 fps. Digital zoom of Video 1. Download video
- Video 3
Disrupting CD44-HA binding with mAb IM7 impairs DC interactions with human lymphatic endothelial cells. Representative spinning disc confocal microscopy time-lapse of monocyte-derived DC labeled with CMFDA green tracker dye, crawling on but not stably adhering to a monolayer of confluent human lymphatic endothelial cell labeled with anti–LYVE-1 (mAb6A) conjugated with DyLight 650 fluorophore (red), after co-incubation with anti-CD44 mAb IM7. Scale bar = 100 μm, time (mins:sec), 20 fps. Video corresponds to the image in Fig 3C and D.Download video
- Video 4
Disrupting CD44-HA binding with mAb BRIC235 impairs DC interactions with human lymphatic endothelial cells. Representative spinning disc confocal microscopy time-lapse of monocyte-derived DC labeled with CMFDA green tracker dye, crawling on but not stably adhering to a monolayer of confluent human lymphatic endothelial cell labeled with anti–LYVE-1 (mAb6A) conjugated with DyLight 650 fluorophore (red), after co-incubation with anti-CD44 mAb BRIC235. Scale bar = 100 μm, time (mins:sec), 20 fps. Video corresponds to the image in Fig 3C and D.Download video
- Video 5
Co-localization of CD44 with the uropod marker ICAM3 in monocyte-derived DCs crawling on human lymphatic endothelial cell monolayer. Representative time-lapse spinning disc confocal microscopy of monocyte-derived DC labeled with non-function blocking anti-CD44 mAb F10.44.2 (FITC-conjugated, green) and anti-ICAM3 (APC-conjugated, red), crawling on a monolayer of confluent human lymphatic endothelial cell. Scale bar = 100 μm, time h:min, 2 fps. Video corresponds to the image in Fig 3B and E.Download video
- Video 6
Localization of CD44 in the uropod of adherent monocyte-derived DCs. Representative spinning disc confocal microscopy time-lapse of monocyte-derived DCs labeled with anti-CD44 mAb F10.44.2 (FITC-conjugated, green) and anti-ICAM3 (APC-conjugated, red), crawling on a monolayer of confluent human lymphatic endothelial cell. Scale bar = 20 μm, time h:min, 2 fps. Digital zoom of Video 5.Download video
- Video 7
CD44 deficiency impairs BMDC interactions with mouse lymphatic endothelial cell (mLEC) monolayer. Representative spinning disc confocal microscopy time-lapse of CD44+/+ BMDC (green, CMFDA) adhering and migrating across mLEC monolayer, in contrast to CD44−/− BMDC (red, CMPXT), crawling over but not stably adhering to a monolayer of confluent mLEC. A dragon tail path of CD44−/− BMDC migration is shown (Image J). Scale bar = 10 μm, 5 fps.Download video




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
