



Article Figures & Data
Figures
- Figure 1. Activation of ATFS-1 up-regulates genes from multiple stress response pathways.
To determine the role of ATFS-1 in the activation of genes from different stress response pathways, we activated ATFS-1 by mildly impairing mitochondrial function through a mutation in nuo-6 (blue bars) and then examined the effect of disrupting atfs-1 using an atfs-1 deletion mutant atfs-1(gk3094) (purple bars). We also examined the expression of these genes in two constitutively active atfs-1 mutants, atfs-1(et15) and atfs-1(et17) (green bars). (A, B, C, D, E, F, G, H) Target genes from the mitochondrial unfolded protein response (A, mitoUPR, hsp-6), the ER unfolded protein response (B, ER-UPR, hsp-4), the cytoplasmic unfolded protein response (C, Cyto-UPR, hsp-16.2), the DAF-16–mediated stress response (D, sod-3), SKN-1–mediated oxidative stress response (E, gst-4), HIF-1–mediated hypoxia response (F, nhr-57), p38-mediated innate immune pathway (G, Y9C9A.8), and antioxidant defense (H, trx-2) were measured. Target genes from the mitoUPR, DAF-16–mediated stress response, SKN-1–mediated oxidative stress response, HIF-1–mediated hypoxia response, p38-mediated innate immune pathway, and antioxidant defense are all significantly up-regulated in nuo-6 mutants in an ATFS-1–dependent manner. Target genes from the mitoUPR, DAF-16–mediated stress response, HIF-1–mediated hypoxia response, p38-mediated innate immune pathway, and antioxidant defense are also up-regulated in at least one of the constitutively activated atfs-1 mutants. In contrast, activation of ATFS-1 by nuo-6 mutation or atfs-1 gain-of-function mutations did not significantly affect target gene expression for the ER-UPR or the Cyto-UPR. atfs-1(gk3094) is a loss-of-function deletion mutant. atfs-1(et15) and atfs-1(et17) are constitutively active gain-of-function mutants. A full list of genes that are up-regulated by ATFS-1 activation can be found in Table S2. Data information: Error bars indicate SEM. **P < 0.01, ***P < 0.001. Statistical analysis was performed using a one-way ANOVA with the Bonferroni post hoc test. The number of replicates and statistical analysis can be found in Table S6.
- Figure 2. Constitutive activation of ATFS-1 results in up-regulation of genes from multiple stress response pathways.
(A, B, C, D, E, F, G, H) Genes that are up-regulated by constitutive activation of ATFS-1 were compared with previously published lists of genes involved in different stress response pathways, including the mitochondrial unfolded protein response (A, mitoUPR), the ER unfolded protein response (B, ER-UPR), the cytoplasmic unfolded protein response (C, Cyto-UPR), the DAF-16–mediated stress response (D), the SKN-1–mediated oxidative stress response (E), the HIF-1–mediated hypoxia response (F), the p38-mediated innate immune response (G), and antioxidant genes (H). In every case, there was a significant degree of overlap ranging from 22 to 51%. Grey circles indicate genes that are up-regulated by activation of the stress response pathway indicated. Turquoise circles indicate genes that are up-regulated in the atfs-1(et15) constitutively active gain-of-function mutant. The numbers inside the circles show how many genes are up-regulated. The percentage overlap is the number of overlapping genes as a percentage of the number of genes up-regulated by the stress response pathway. P-values indicate the significance of the difference between the observed number of overlapping genes between the two gene sets, and the expected number of overlapping genes if the genes were picked at random. Panel (I) shows an inclusive UpSetR plot displaying the overlap between up-regulated genes associated with each stress response pathway. Vertical bars indicate the number of genes in common (overlap) between gene sets indicated by the dots below. Horizontal black bars indicate the number of genes within each gene set. mitoUPR, mitochondrial unfolded protein response; ER-UPR, endoplasmic reticulum unfolded protein response; Cyto-UPR, cytoplasmic unfolded protein response; DAF-16, DAF-16–mediated stress response pathway; SKN-1, SKN-1–mediated oxidative stress response pathway; HIF-1, HIF-1–mediated hypoxia response pathway; innate immunity, p38-mediated innate immunity pathway; antioxidant, antioxidant genes. Stress pathway gene lists and sources can be found in Table S3. Lists of genes common to multiple stress response pathways can be found in Table S4.
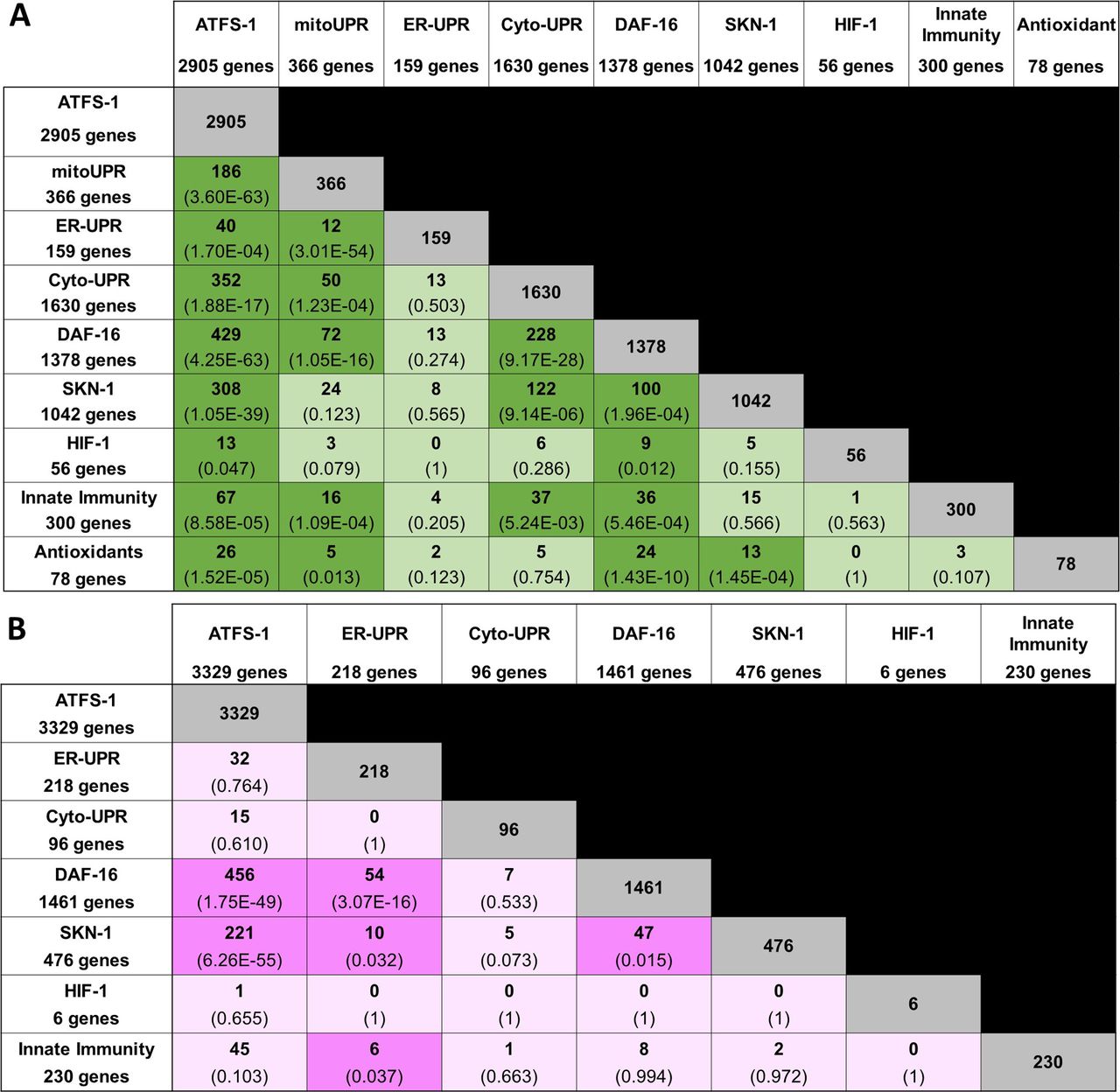
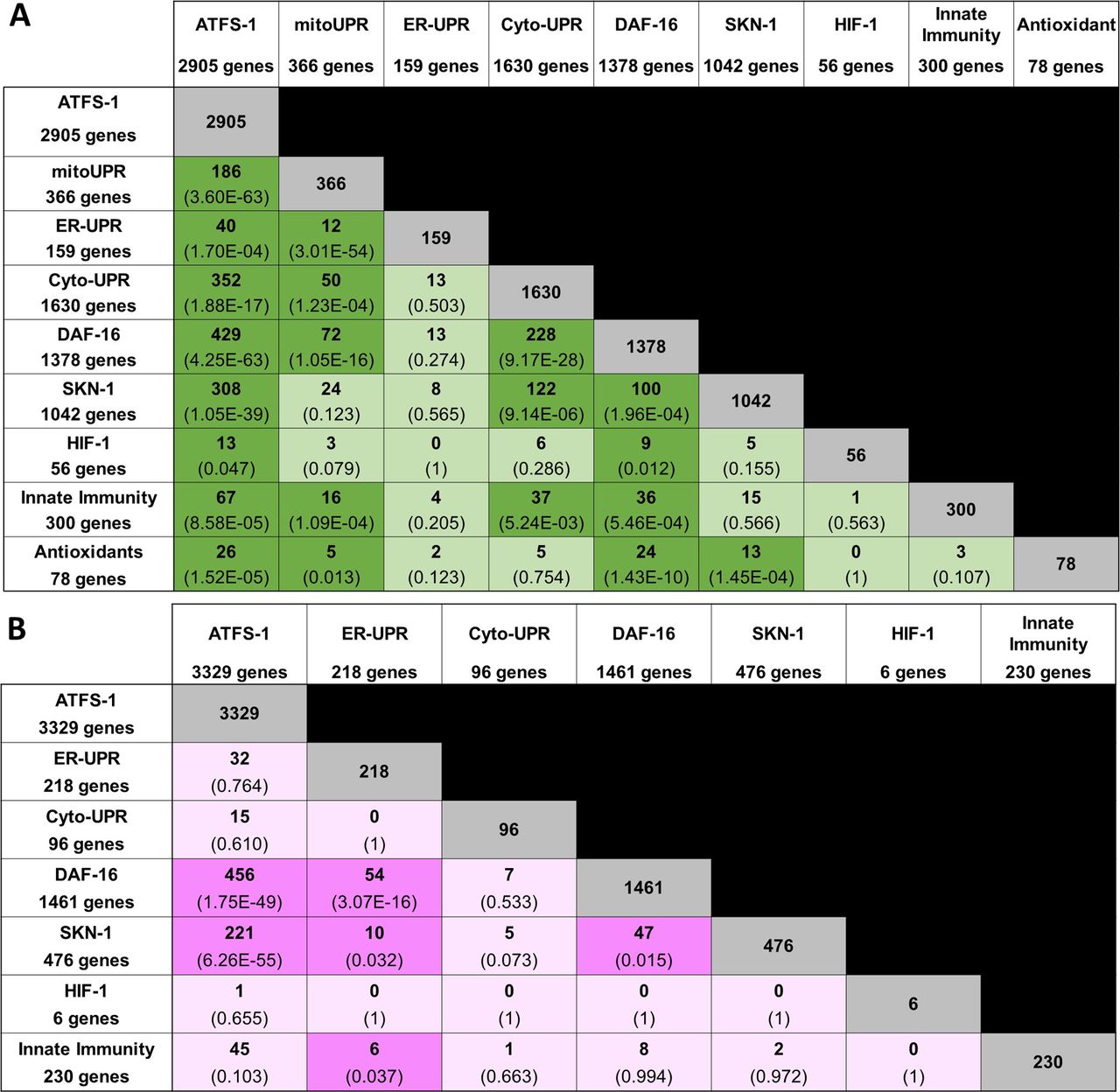
- Figure S1. Overlap in gene expression between different stress response pathways.
(A, B) Panel (A) shows up-regulated genes; panel (B) shows down-regulated genes. The number of overlapping genes is indicated in each box along with the statistical significance of this overlap (P-value in brackets). The size of each gene set is indicated in the first column and row. ATFS-1, genes up-regulated by activation of ATFS-1 (genes up-regulated in atfs-1(et15) mutant); mitoUPR, mitochondrial unfolded protein response (genes up-regulated by spg-7 RNAi in an atfs-1–dependent manner); ER-UPR, endoplasmic reticulum unfolded protein response; Cyto-UPR, cytoplasmic unfolded protein response; DAF-16, DAF-16–mediated stress response pathway; SKN-1, SKN-1–mediated oxidative stress response pathway; HIF-1, HIF-1–mediated hypoxia response pathway; innate immunity, p38-mediated innate immunity pathway; antioxidant, antioxidant genes. Stress pathway gene lists and sources can be found in Table S3. Lists of genes common to multiple stress response pathways can be found in Table S4.
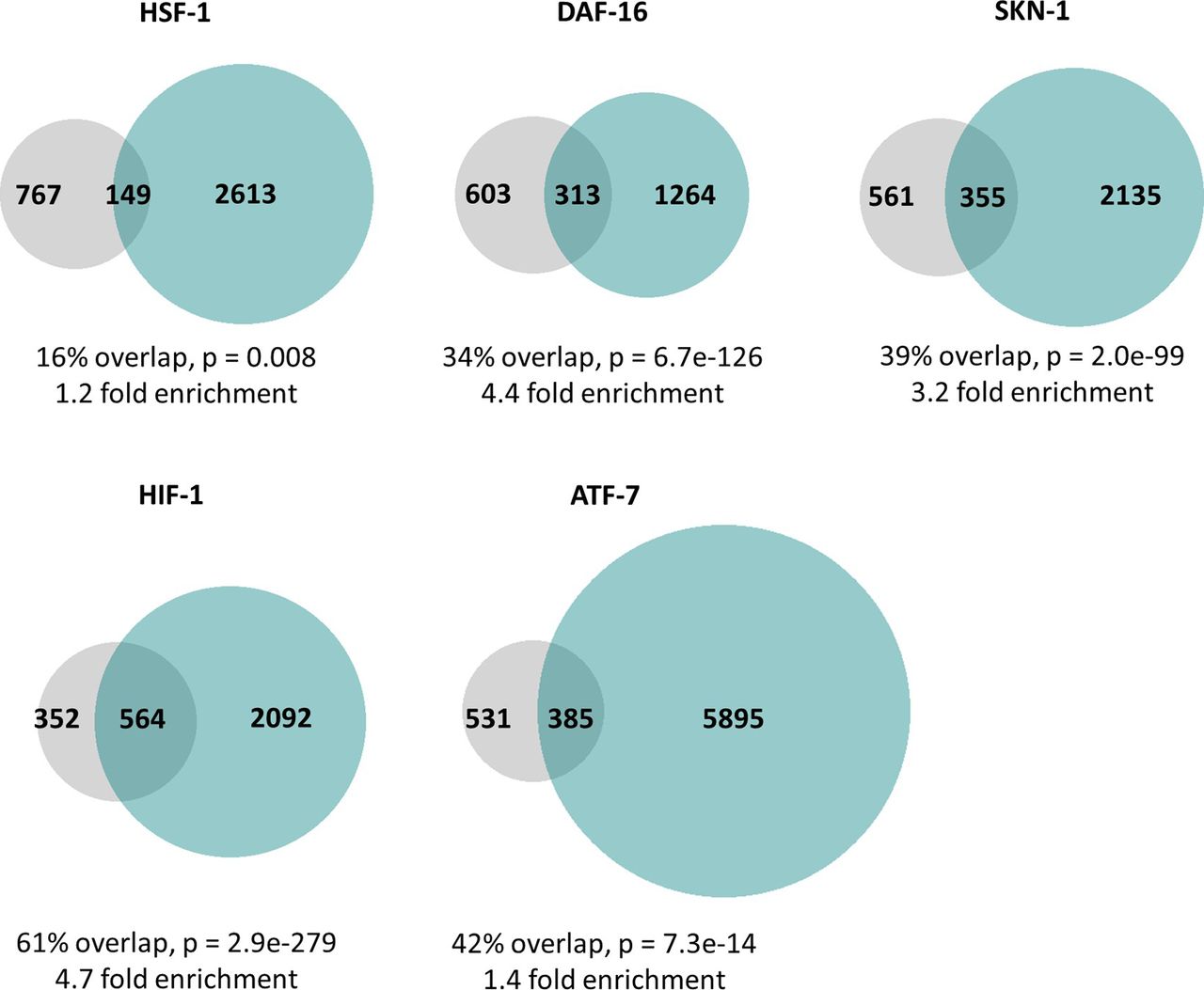
- Figure S2. ATFS-1 can bind to the same promoters as other stress-responsive transcription factors.
In order to better understand the mechanisms underlying the overlap in gene expression changes between constitutively active atfs-1 mutants and other stress response pathways, we used previously published ChIP-seq datasets to determine if ATFS-1 can bind to the same genes other stress-responsive transcription factors including HSF-1 (cytoplasmic unfolded protein response), DAF-16 (DAF-16–mediated stress response), SKN-1 (SKN-1–mediated oxidative stress response), HIF-1 (HIF-1–mediated hypoxia response), and ATF-7 (p38-mediated innate immune response). In every case, we found that there were a set of genes that could be bound by both ATFS-1 and the stress-responsive transcription factor, and the number of genes in this overlapping set was significantly larger than if the genes were picked at random. The degree of enrichment compared with the expected number of genes in the overlap ranged from 1.2-fold to 4.7-fold enrichment. Grey circles indicate genes bound by ATFS-1. Turquoise circles indicate genes bound by the stress-responsive transcription factor indicated. The percentage overlap is the number of overlapping genes as a percentage of the number of genes bound by ATFS-1. P-values indicate the significance of the difference between the observed number of overlapping genes between the two gene sets, and the expected number of overlapping genes if the genes were picked at random.
- Figure 3. ATFS-1 is required for up-regulation of stress response genes after exposure to oxidative stress or bacterial pathogen stress.
To determine the role of ATFS-1 in responding to different types of stress, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to different stressors. (A) Exposure to oxidative stress (4 mM paraquat, 48 h) caused a significant up-regulation of hsp-6, nhr-57, and trx-2 in wild-type worms that was prevented by the disruption of atfs-1. (B) Exposure to bacterial pathogen stress (PA14, 24 h) resulted in an up-regulation of nhr-57, gcs-1 and sod-5 in wild-type worms that was prevented by the atfs-1 deletion. (C) Exposure to heat stress (35°C, 2 h) caused increased expression of ckb-2 and a trend towards increased expression of hsp-16.2 and hsp-4 in both wild-type and atfs-1 worms. (D) Exposure to osmotic stress (300 mM, 24 h) caused an up-regulation of sod-3, gst-4, and Y9C9A.8 in wild-type worms and to a greater magnitude in atfs-1 mutants. (E) Anoxia (24 h) resulted in the up-regulation of hsp-16.2, Y9C9A.8, and dod-22 in both wild-type and atfs-1 worms. (F) Exposing worms to ER stress (5 μg/ml tunicamycin, 24 h) increased the expression of ckb-2 and trended towards increasing the expression of clec-67 in both wild-type and atfs-1 worms. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. The number of replicates and statistical analysis can be found in Table S6.
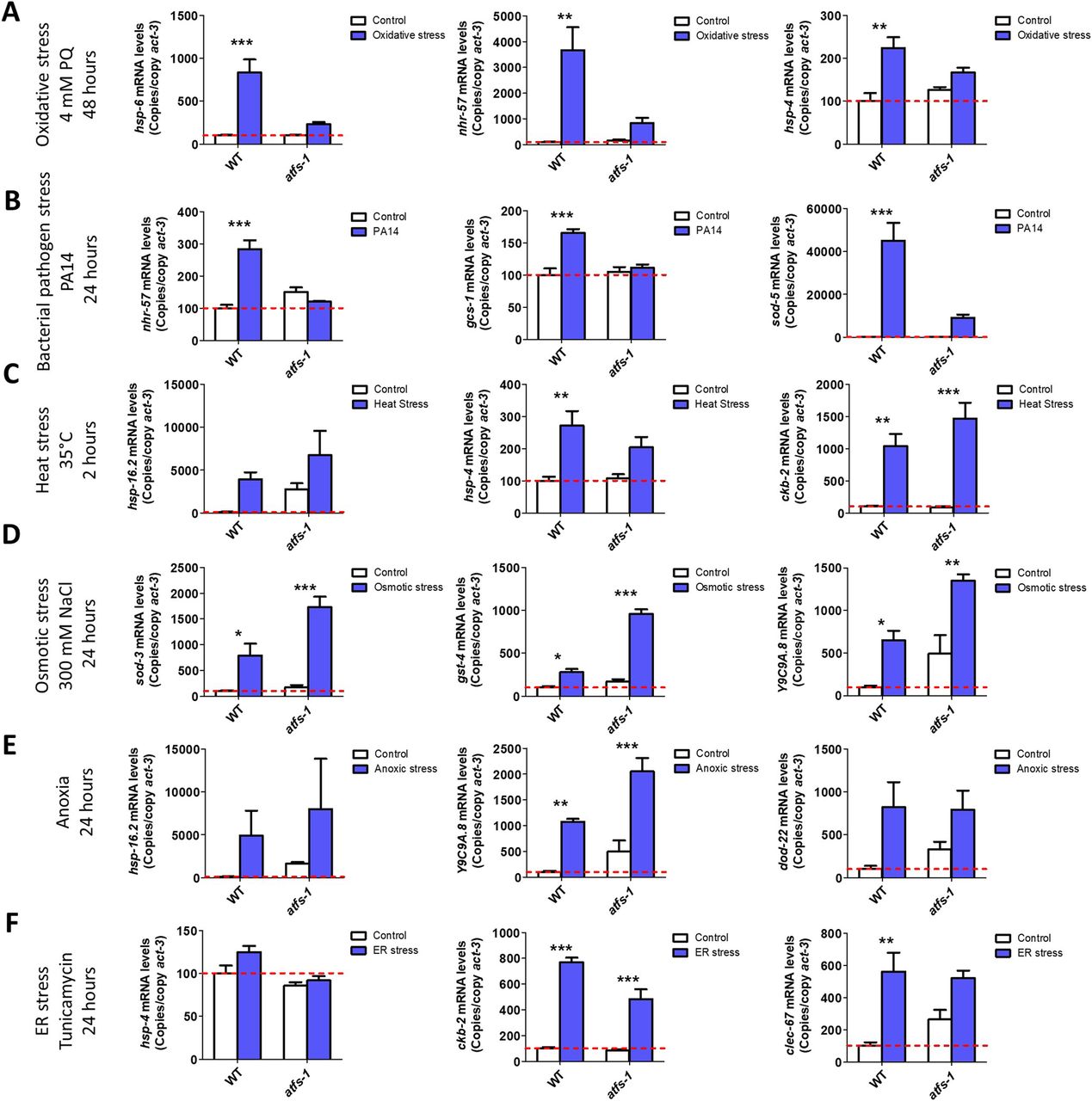
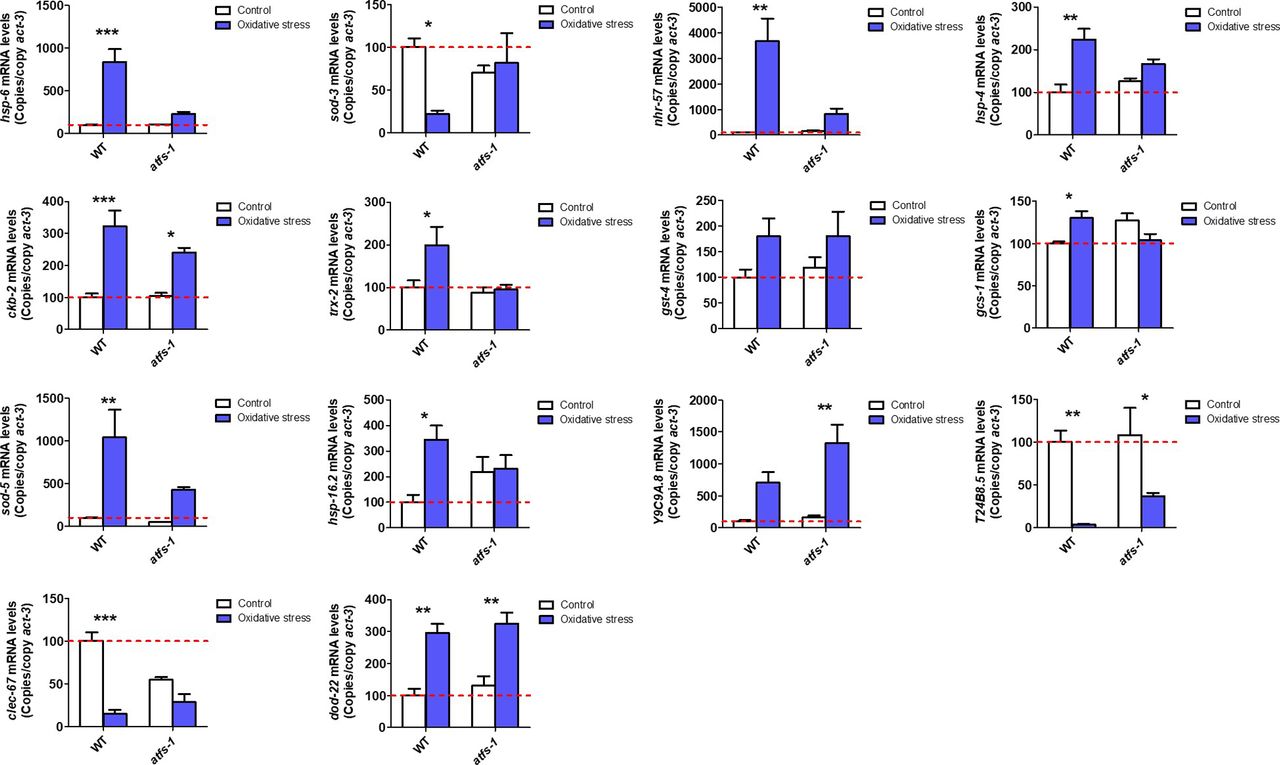
- Figure S3. ATFS-1 is required for up-regulation of stress response genes after exposure to oxidative stress.
To determine the role of ATFS-1 in responding to oxidative stress, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to 4 mM paraquat for 48 h. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. Three biological replicates were performed.
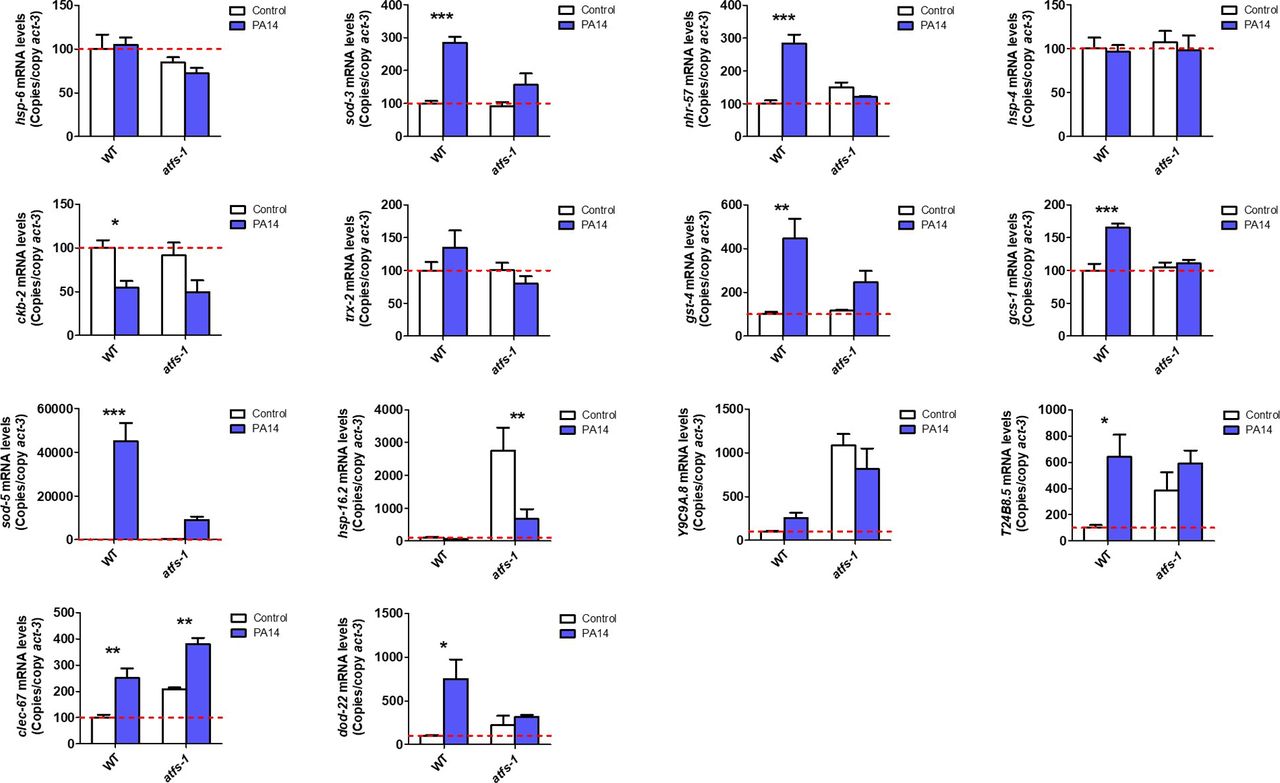
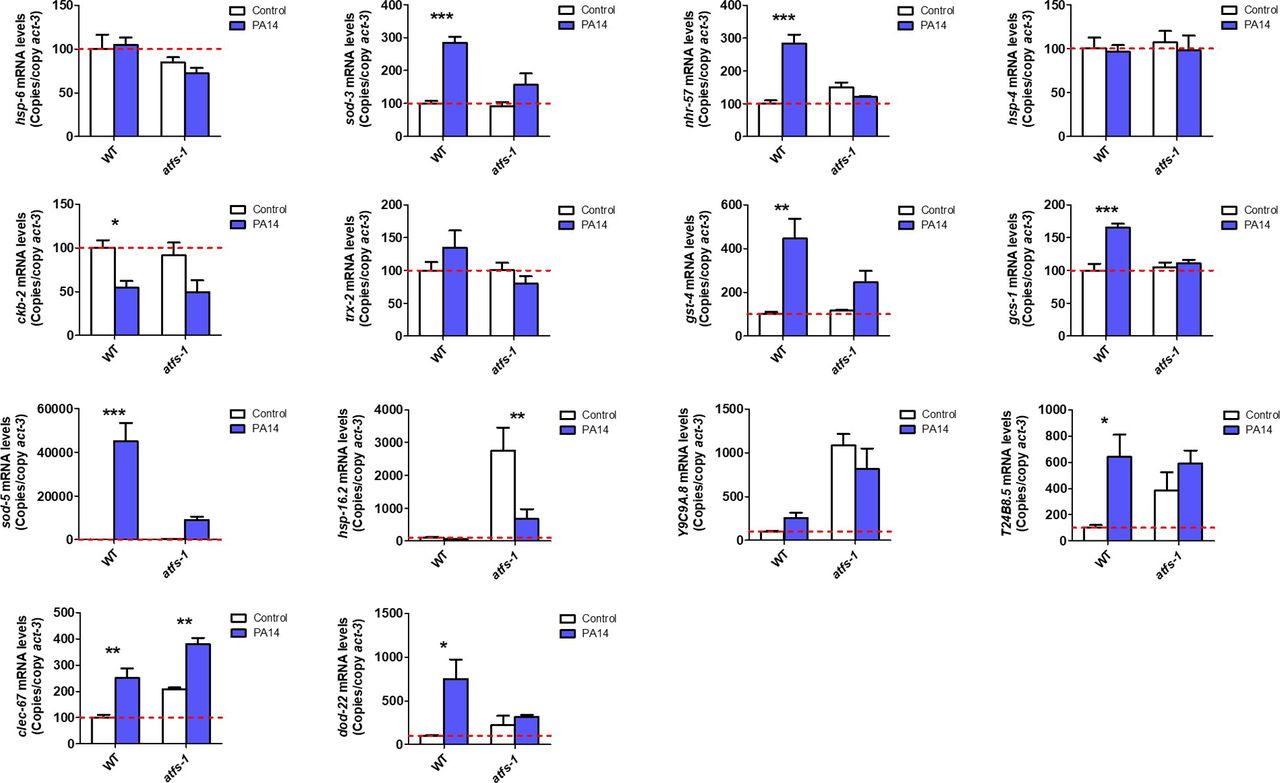
- Figure S4. ATFS-1 is required for up-regulation of stress response genes after exposure to bacterial pathogens.
To determine the role of ATFS-1 in responding to bacterial pathogens, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to Pseudomonas aeruginosa strain PA14 for 4 h. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. Three biological replicates were performed.
- Figure S5. ATFS-1 is not required for up-regulation of stress response genes after exposure to heat stress.
To determine the role of ATFS-1 in responding to heat stress, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to 35°C heat for 2 h followed by 20°C for 4 h. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. Three biological replicates were performed.
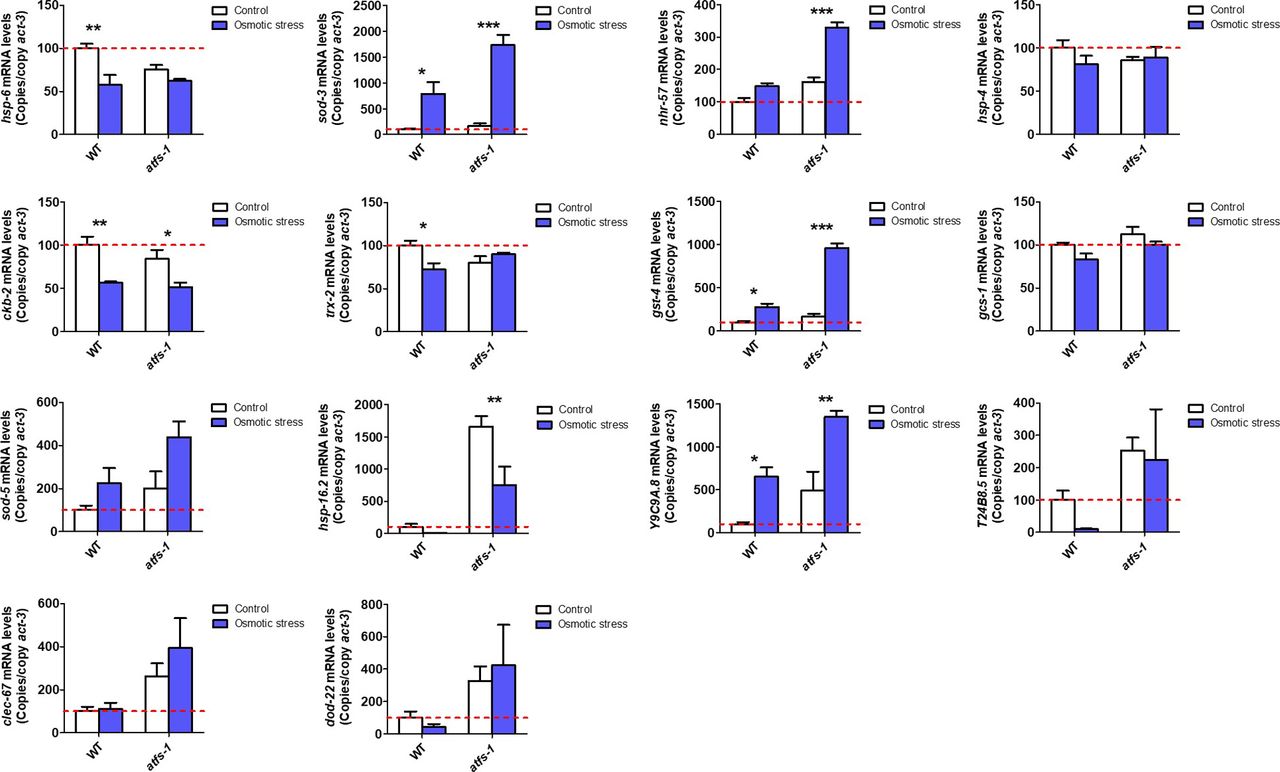
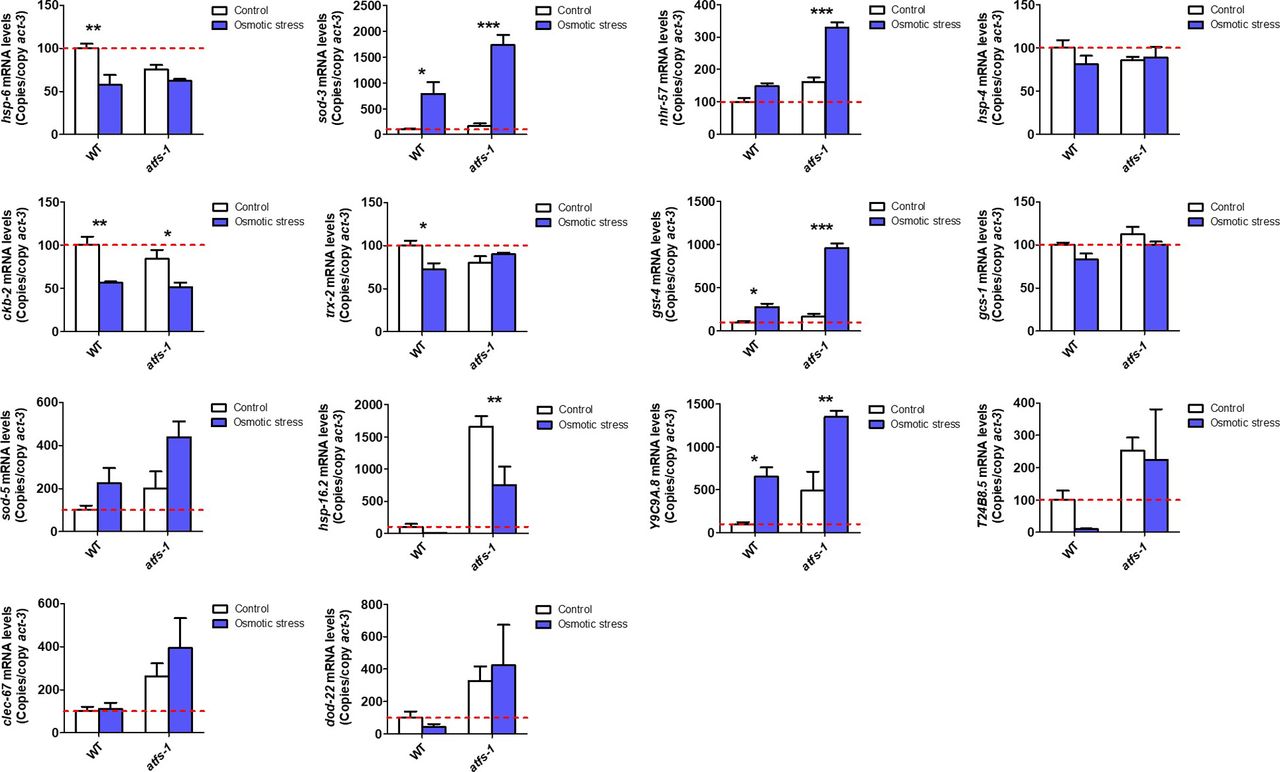
- Figure S6. ATFS-1 is not required for up-regulation of stress response genes after exposure to osmotic stress.
To determine the role of ATFS-1 in responding to osmotic stress, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to 300 mM NaCl heat for 24 h. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. Three biological replicates were performed.
- Figure S7. ATFS-1 is not required for up-regulation of stress response genes after exposure to anoxic stress.
To determine the role of ATFS-1 in responding to anoxic stress, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to complete anoxia for 24 h followed by a 4-h recovery at normoxia. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. Three biological replicates were performed.
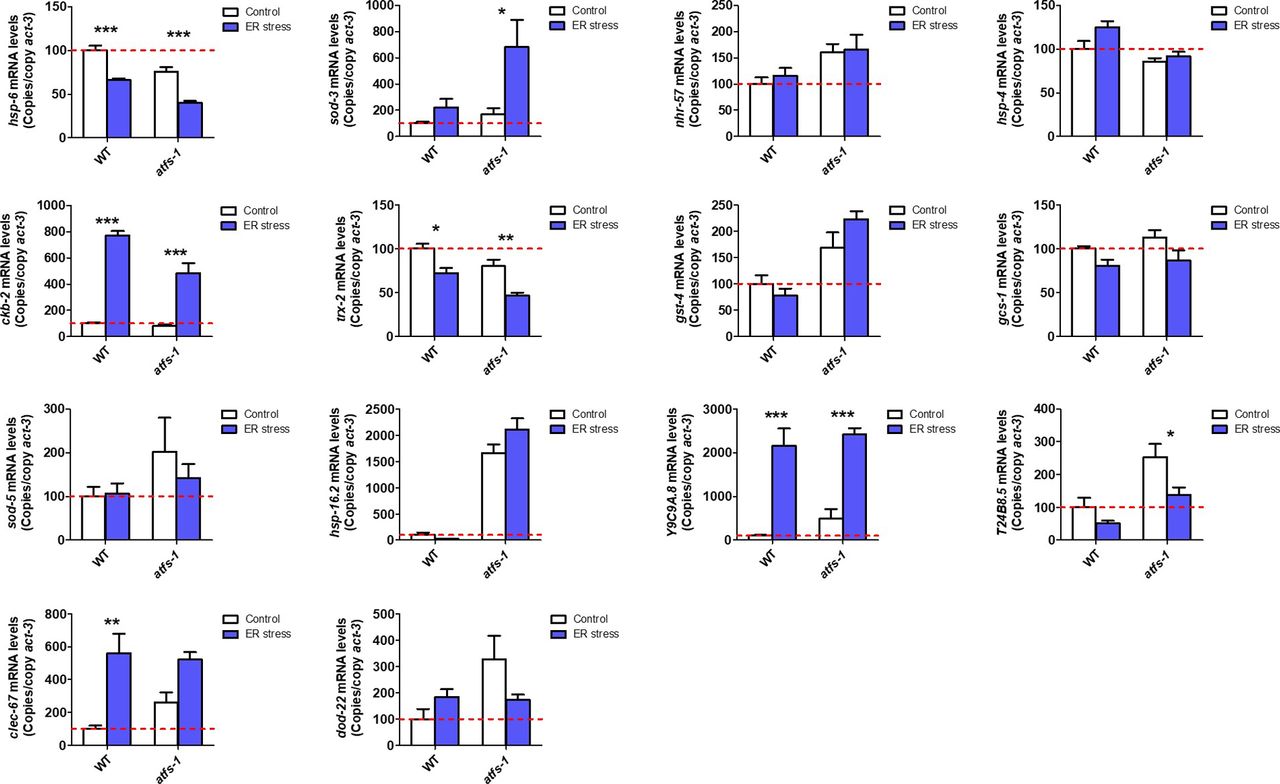
- Figure S8. ATFS-1 is not required for up-regulation of stress response genes after exposure to ER stress.
To determine the role of ATFS-1 in responding to ER stress, we compared the up-regulation of stress response genes in wild-type and atfs-1(gk3094) loss-of-function deletion mutants after exposure to 5 μg/ml tunicamycin for 24 h. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis was performed using a two-way ANOVA with a Bonferroni post hoc test. Three biological replicates were performed.
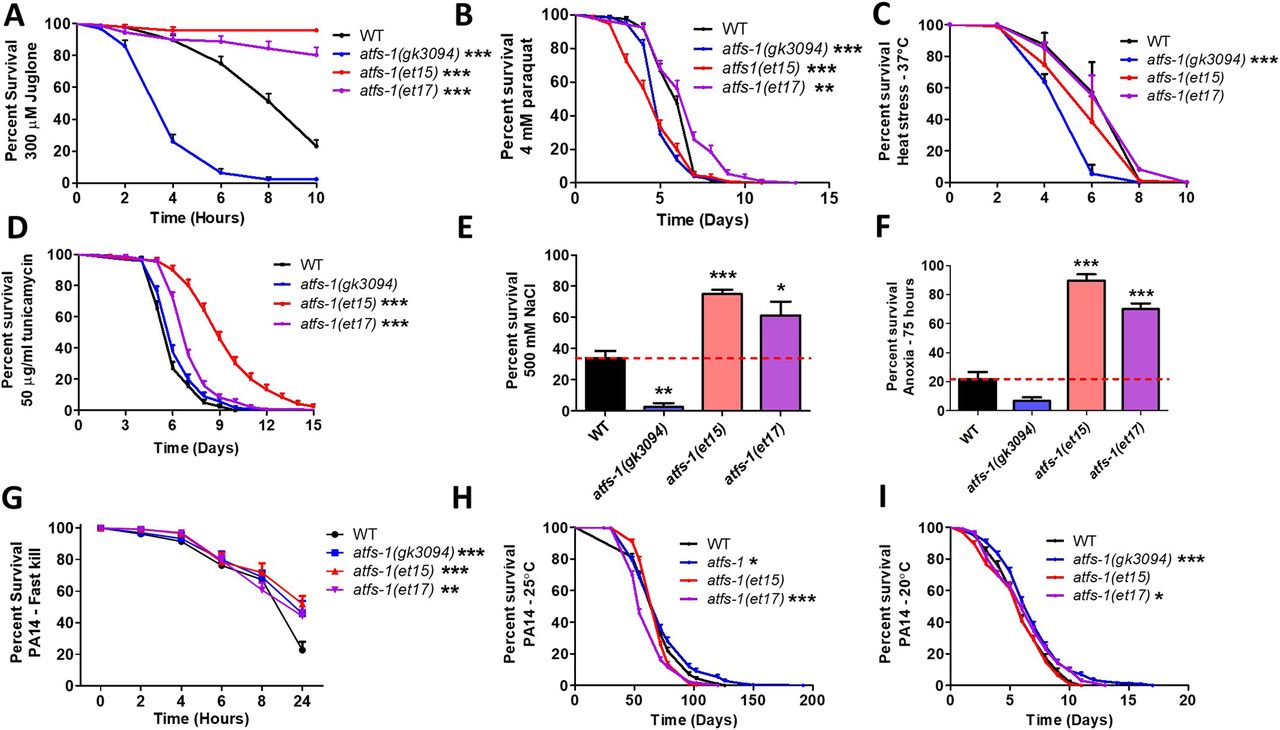
- Figure 4. Constitutive activation of ATFS-1 increases resistance to multiple external stressors.
To determine the role of ATFS-1 in resistance to stress, the stress resistance of an atfs-1 loss-of-function mutants (atfs-1(gk3094)) and two constitutively active atfs-1 gain-of-function mutants (atfs-1(et15), atfs-1(et17)) was compared with wild-type worms. (A) Activation of ATFS-1 enhanced resistance to acute oxidative stress (300 μM juglone), whereas disruption of atfs-1 markedly decreased resistance to acute oxidative stress. (B) Disruption of atfs-1 decreased resistance to chronic oxidative stress (4 mM paraquat). atfs-1(et17) mutants showed increased resistance to chronic oxidative stress, whereas atfs-1(et15) mutants had decreased resistance. (C) Resistance to heat stress (37°C) was not enhanced by activation of ATFS-1, whereas disruption of atfs-1 decreased heat stress resistance. (D) Constitutive activation of ATFS-1 increased resistance to ER stress (50 μM tunicamycin), whereas disruption of atfs-1 had no effect. (E) Activation of ATFS-1 increased resistance to osmotic stress (500 mM NaCl), whereas disruption of atfs-1 decreased osmotic stress resistance. (F) Constitutively active atfs-1 mutants show increased resistance to anoxia (75 h), whereas atfs-1 deletion mutants exhibit a trend towards decreased anoxia resistance. (G) Activation of ATFS-1 increased resistance to Pseudomonas aeruginosa toxin in a fast kill assay. A slow kill assay in which worms die from internal accumulation of P. aeruginosa was performed according to two established protocols. (H) At 25°C, atfs-1(et17) mutants showed a small decrease in resistance to bacterial pathogens (PA14), wheras atfs-1(gk3094) mutants showed a small increase in resistance. (I) At 20°C, both atfs-1(et17) and atfs-1(gk3094) mutants exhibited a small increase in resistance to bacterial pathogens. Data for WT and atfs-1(gk3094) in panel (I) are from Campos et al (2021) as these strains were used as controls for two separate experiments that were performed at the same time. Data information: Error bars indicate SEM. *P < 0.05, **P < 0.01, ***P < 0.001. Statistical analysis for panels (A, B, D, H, I) were performed using the log-rank test. Statistical analysis for panels C and G were performed using a two-way ANOVA with Bonferroni post hoc test. Statistical analysis for panels (E, F) was performed using a one-way ANOVA with Bonferroni post hoc test. The number of replicates, N, and statistical analysis can be found in Table S6.
- Figure 5. Multiple long-lived mutants from different pathways of lifespan extension show up-regulation of ATFS-1–dependent genes.
To determine the extent to which long-lived genetic mutants from different pathways of lifespan extension show differential expression of ATFS-1 target genes, we compared genes that are up-regulated in nine different long-lived mutants to a published list of spg-7 RNAi-up-regulated, ATFS-1–dependent target genes (Nargund et al, 2012). clk-1, isp-1, nuo-6, sod-2, daf-2, glp-1, and ife-2 worms all show a highly significant degree of overlap with genes up-regulated by spg-7 RNAi in an ATFS-1–dependent manner. The grey circles represent the 366 genes that are up-regulated by spg-7 RNAi in an ATFS-1–dependent manner. Turquoise circles are genes that are significantly up-regulated in the indicated long-lived mutant based upon our RNA sequencing data. The number of unique and overlapping genes is indicated. Percent overlap is calculated as the number of genes in common between the two gene sets divided by the total number of genes that are up-regulated by spg-7 RNAi in an ATFS-1–dependent manner. Enrichment is calculated as the number of overlapping genes observed divided by the number of overlapping genes predicted if genes were chosen randomly. P-values indicate the significance of the difference between the observed number of overlapping genes between the two gene sets, and the expected number of overlapping genes if the genes were picked at random.
- Figure S9. Multiple long-lived mutants from different pathways of lifespan extension show up-regulation of ATFS-1–dependent genes.
To determine the extent to which long-lived genetic mutants from different pathways of lifespan extension show differential expression of ATFS-1 target genes, we compared genes that are up-regulated in nine different long-lived mutants with genes up-regulated in a constitutively active atfs-1 mutant (et15). All of the long-lived mutant worms, except for osm-5, show a highly significant degree of overlap with the constitutively active atfs-1 mutant. The grey circles represent genes that are significantly up-regulated in the constitutively active atfs-1(et15) mutant. Turquoise circles are genes that are significantly up-regulated in the long-lived mutant indicated. The number of unique and overlapping genes is indicated. Overlap is calculated as the number of genes in common between the two gene sets divided by the smaller gene set. Enrichment is calculated as the number of overlapping genes observed divided by the number of overlapping genes predicted if genes were chosen randomly. P-values indicate the significance of the difference between the observed number of overlapping genes between the two gene sets, and the expected number of overlapping genes if the genes were picked at random.
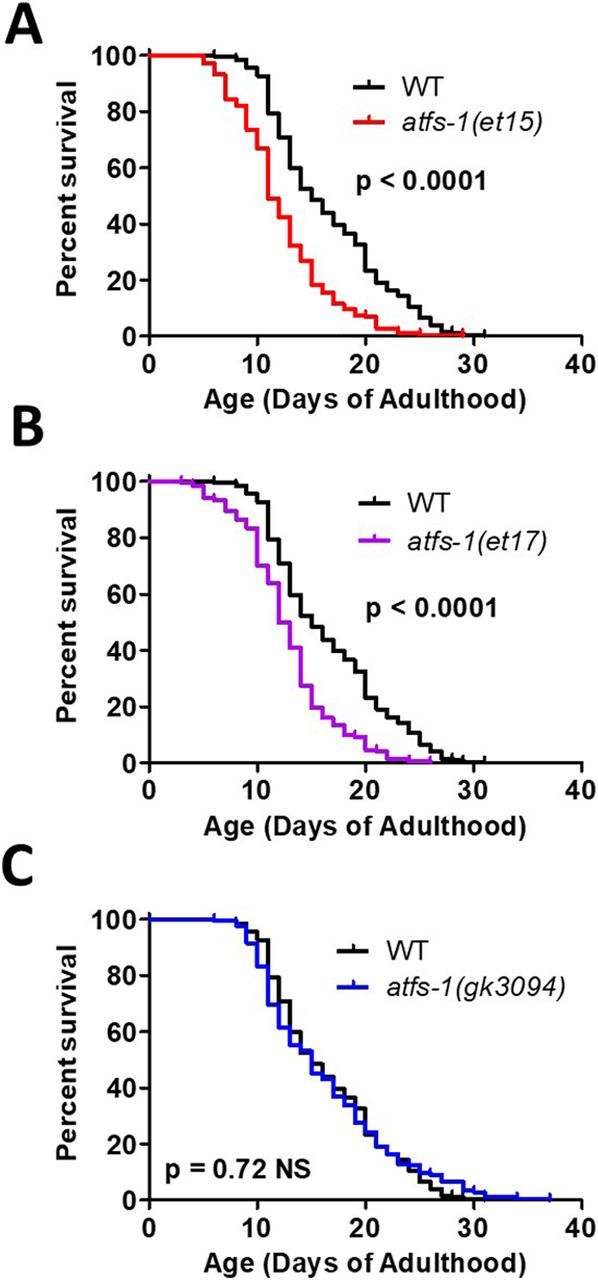
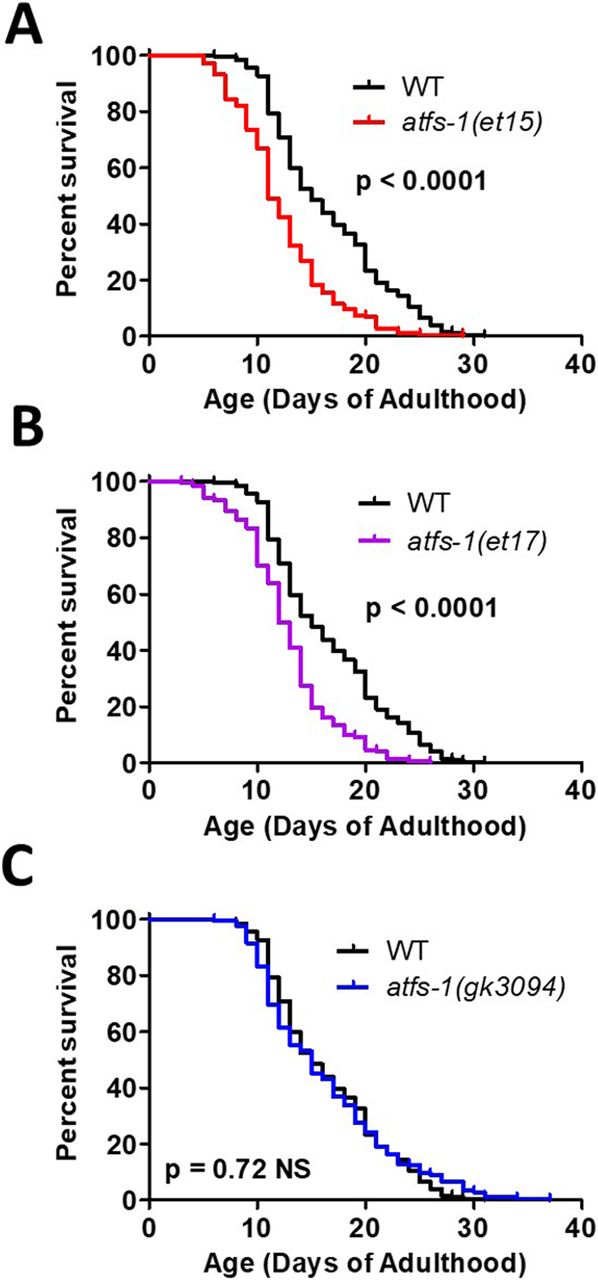
- Figure 6. Activation of ATFS-1 does not increase lifespan.
To determine the effect of ATFS-1 on aging, we quantified the lifespan of an atfs-1 deletion mutant and two constitutively active atfs-1 mutants. (A, B) Both constitutively active atfs-1 mutants, et15 and et17, have a significantly decreased lifespan compared with wild-type worms. (C) Disruption of atfs-1 does not affect lifespan compared with wild-type worms. atfs-1(gk3094) is a loss of function mutant resulting from a deletion. atfs-1(et15) and atfs-1(et17) are constitutively active gain-of-function mutants. Data information: Statistical analysis was performed using the log-rank test. Statistical analysis, number of replicates, N, and raw lifespan data are available in Table S6.









Supplementary Materials




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
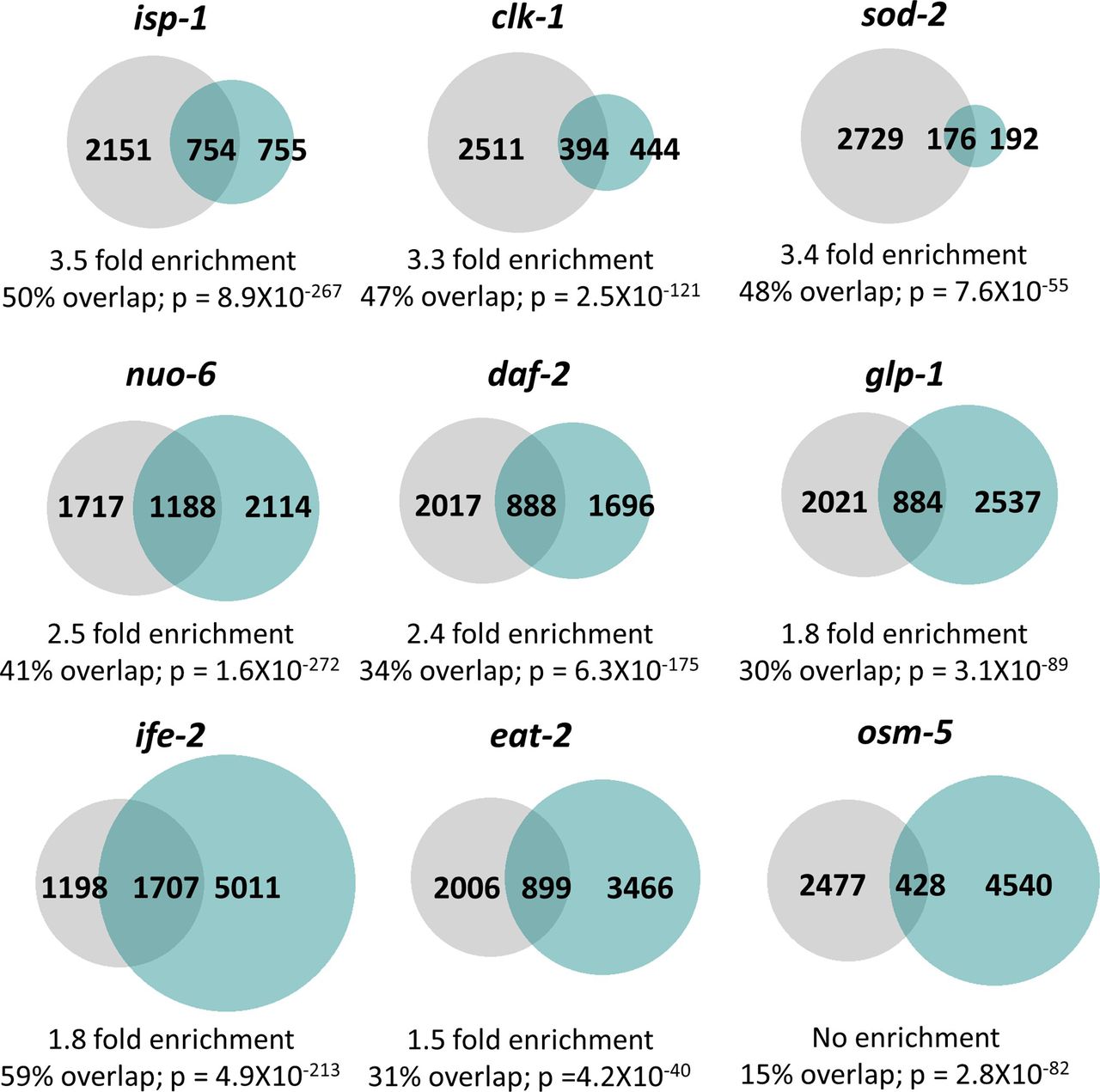{kind=link}
{kind=link}

In this Issue
Related Articles
Cited By...
- No citing articles found.