



Article Figures & Data
Figures
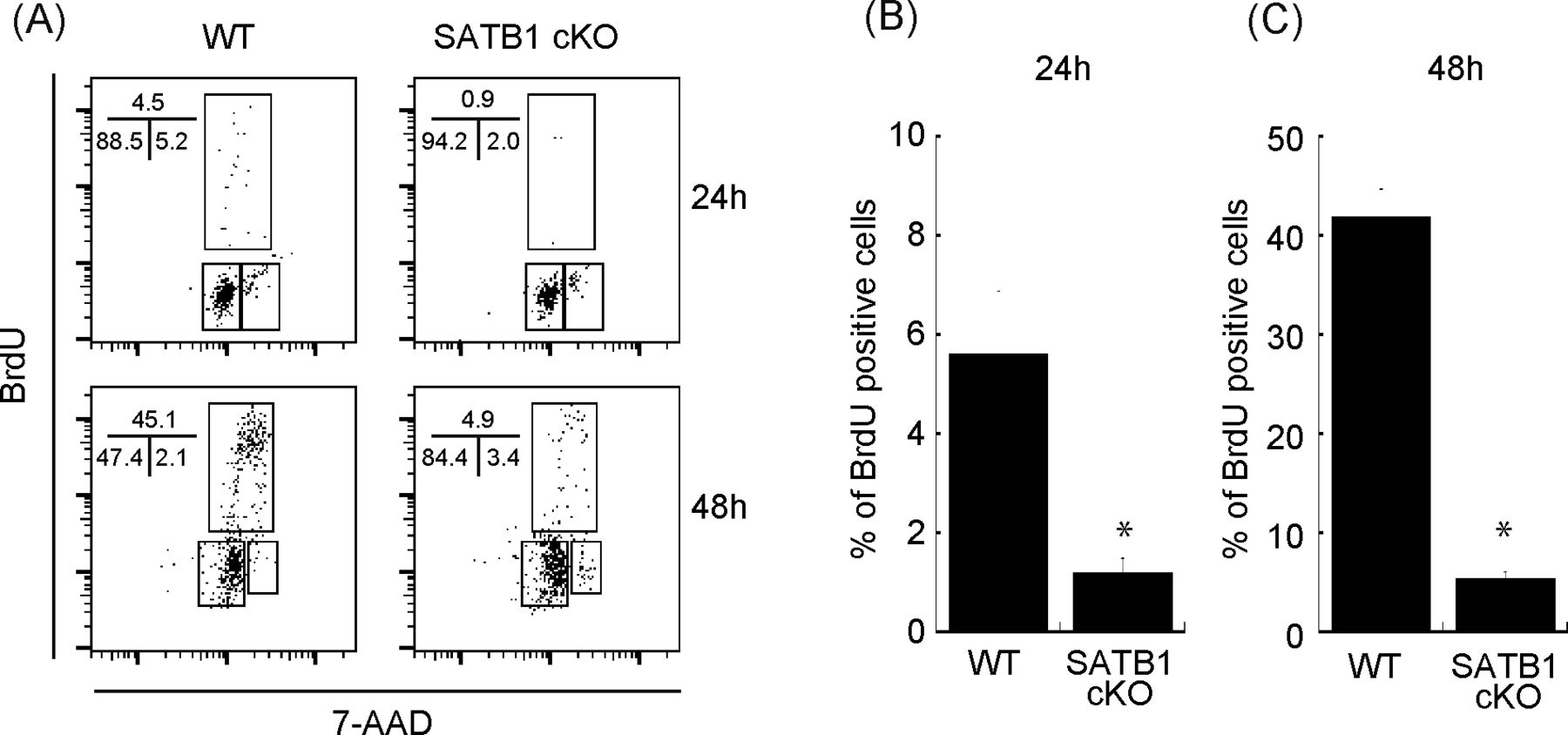
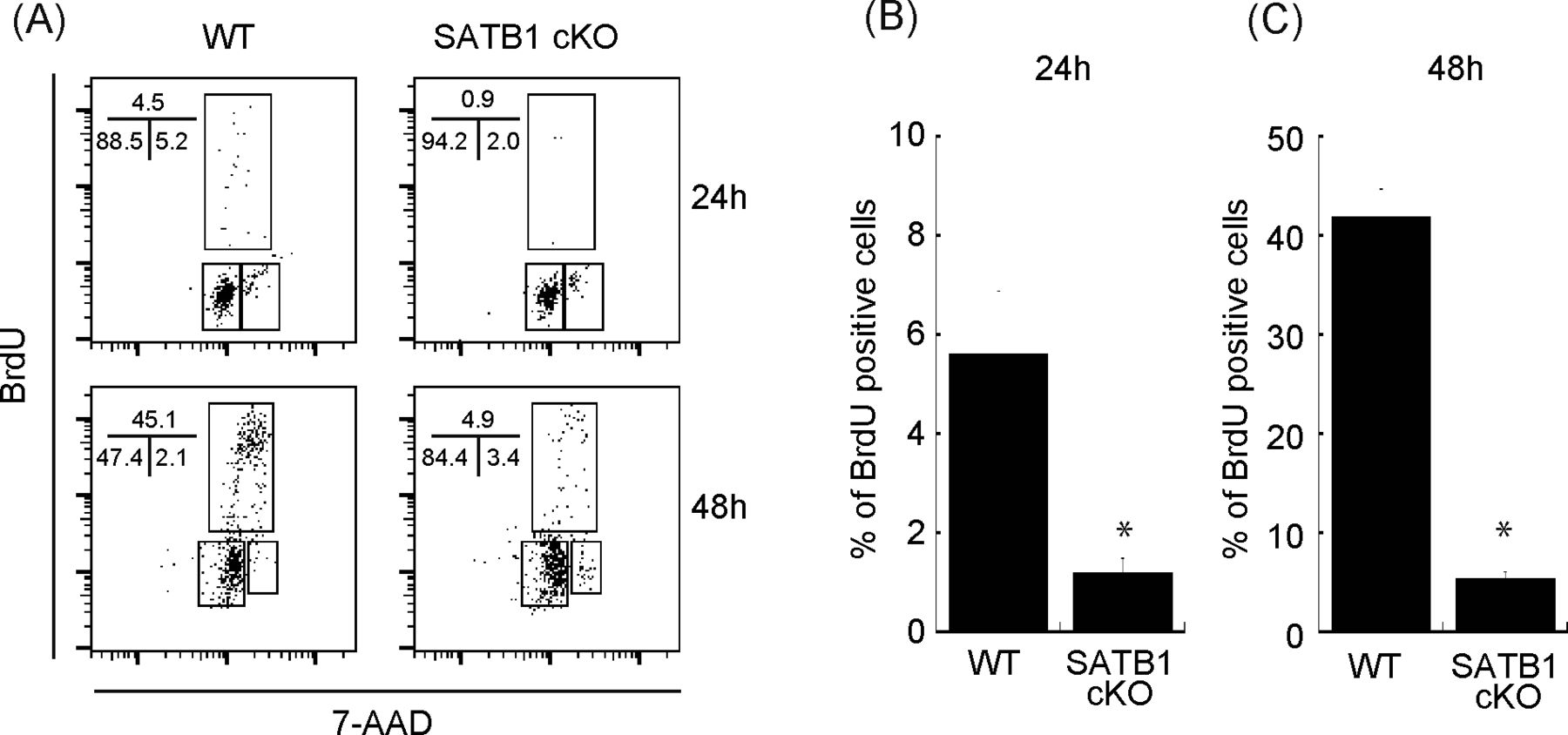
- Figure S1. Cell cycle analysis of CD4 T cells from WT and SATB1cKO mice.
Sorted naïve CD4 T cells were stimulated with anti-CD3 and anti-CD28 antibodies for 24 and 48 h. BrdU was added to the in vitro culture for the last 30 min of incubation. (A) The cells were then fixed and stained with anti-BrdU antibody and 7-amino-actinomycin and subjected to FACS analysis (A). The percentage of cells in each cell cycle phase is shown. Data are representative for three independent experiments. (B, C) The quantification of BrdU-positive cells is shown on the right-hand side (B, C).
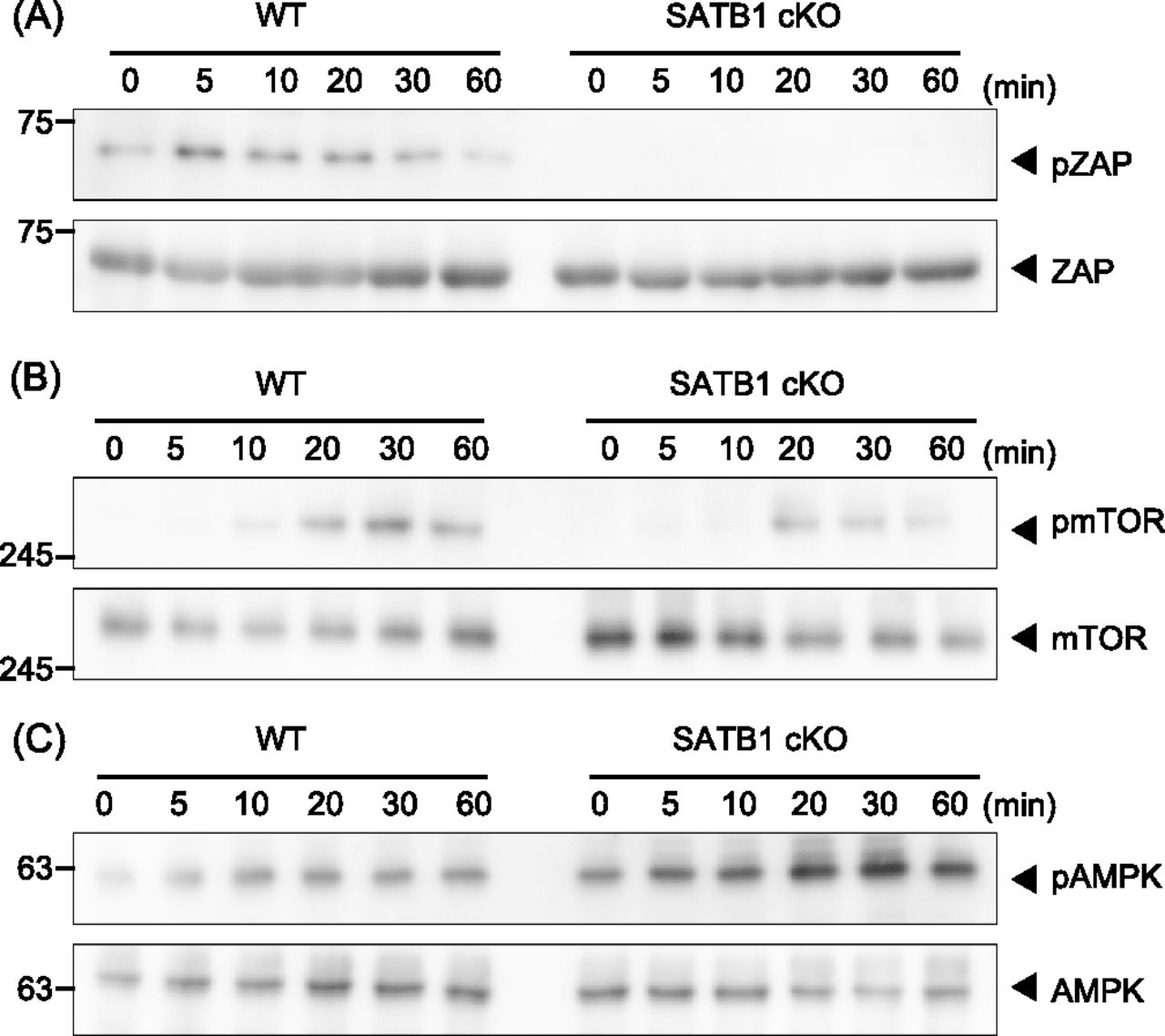
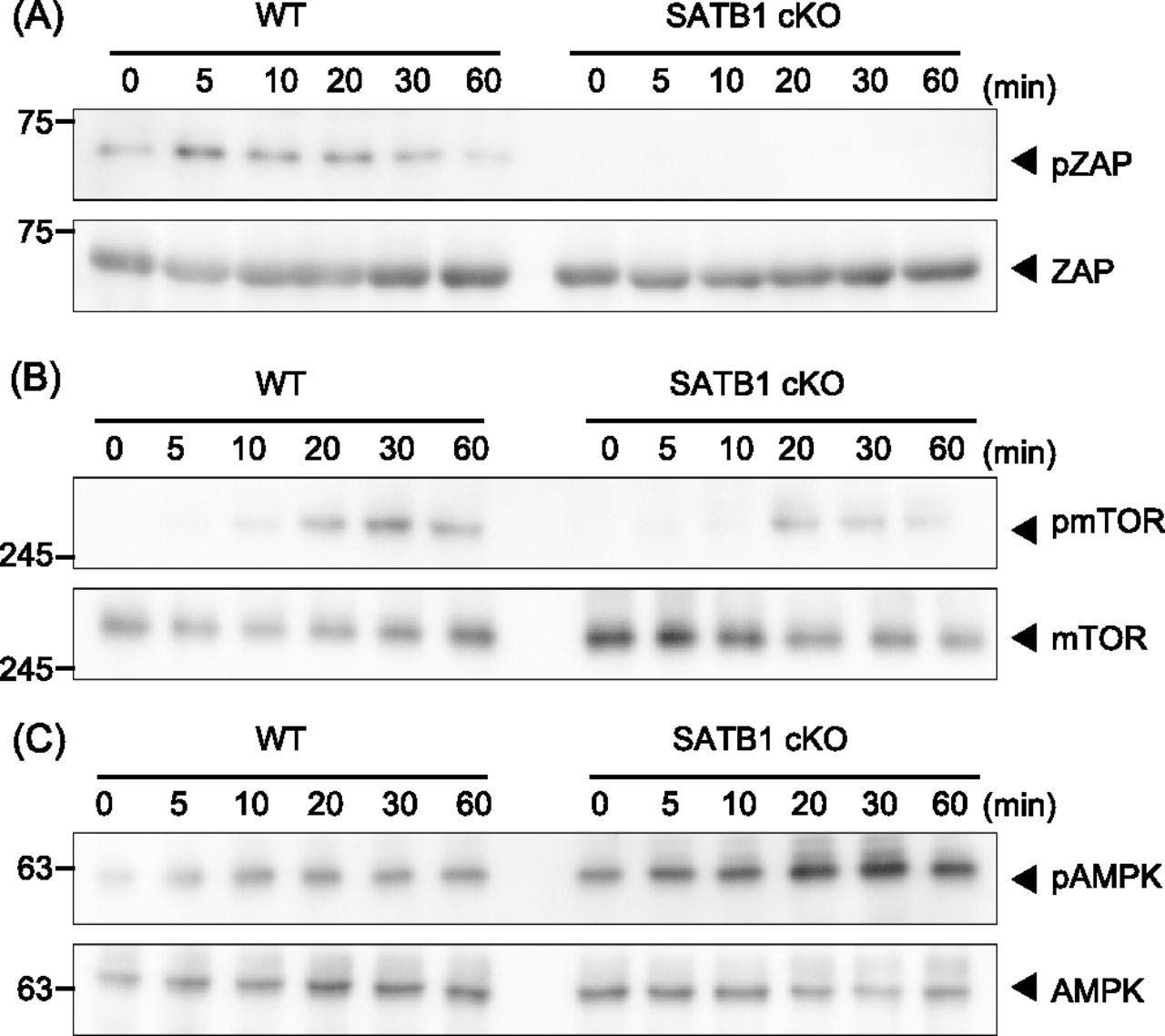
- Figure 1. SATB1 is required for T-cell activation.
Signaling molecule phosphorylation was examined in naïve CD4 T cells from WT and SATB1cKO mice. (A, B, C) T cells were stimulated with anti-CD3 and anti-CD28 antibodies for the indicated time periods, lysed, analyzed by SDS–PAGE, and immunoblotted with anti-ZAP70, anti-phospho-ZAP70 (A), anti-mammalian target of rapamycin, anti-phospho-mammalian target of rapamycin (B), anti-AMPK, and anti-phospho-AMPK (C). Data for one experiment are representative of three independent experiments.
- Figure S2. Signaling molecule phosphorylation examined in naïve CD4 T cells from WT, SATB1cKO, and SATB1cKO-TFAMTg mice.
T cells were stimulated with anti-CD3 and anti-CD28 antibodies for the indicated periods, lysed, analyzed by SDS–PAGE, and immunoblotted with antibodies against p70S6 and phospho-p70S6 kinases. Data for one experiment representative of three independent experiments are shown.
- Figure 2. SATB1-TFAM axis is required for mitochondrial homeostasis.
(A) CD4 T cells were suspended in ATP assay buffer and incubated for 1 min at RT. Light intensity was indicative of intracellular ATP content (Lu-ATP). (B) Oxidative phosphorylation complex I enzyme activity was measured. Lysates were incubated on the plate for 3 h to capture complex I. Enzyme activity was determined by NADH oxidation to NAD+ and simultaneous dye reduction. Dye absorbance was measured with a plate reader at 450 nm. (C) Levels of mtROS in CD4 T cells from WT, and SATB1cKO mice in the control and after TCR activation were measured by DCFDA staining and flow cytometry. (D) Quantification of mean fluorescence of DCFDA. (E) Representative MitoTracker staining histograms. WT and SATB1cKO T cells in the control and after TCR activation were stained with MitoTracker and analyzed by a flow cytometer. (F) Quantification of the mean fluorescence of MitoTracker.*P < 0.01 versus WT. N = 5. Data are shown as the means ± SD.
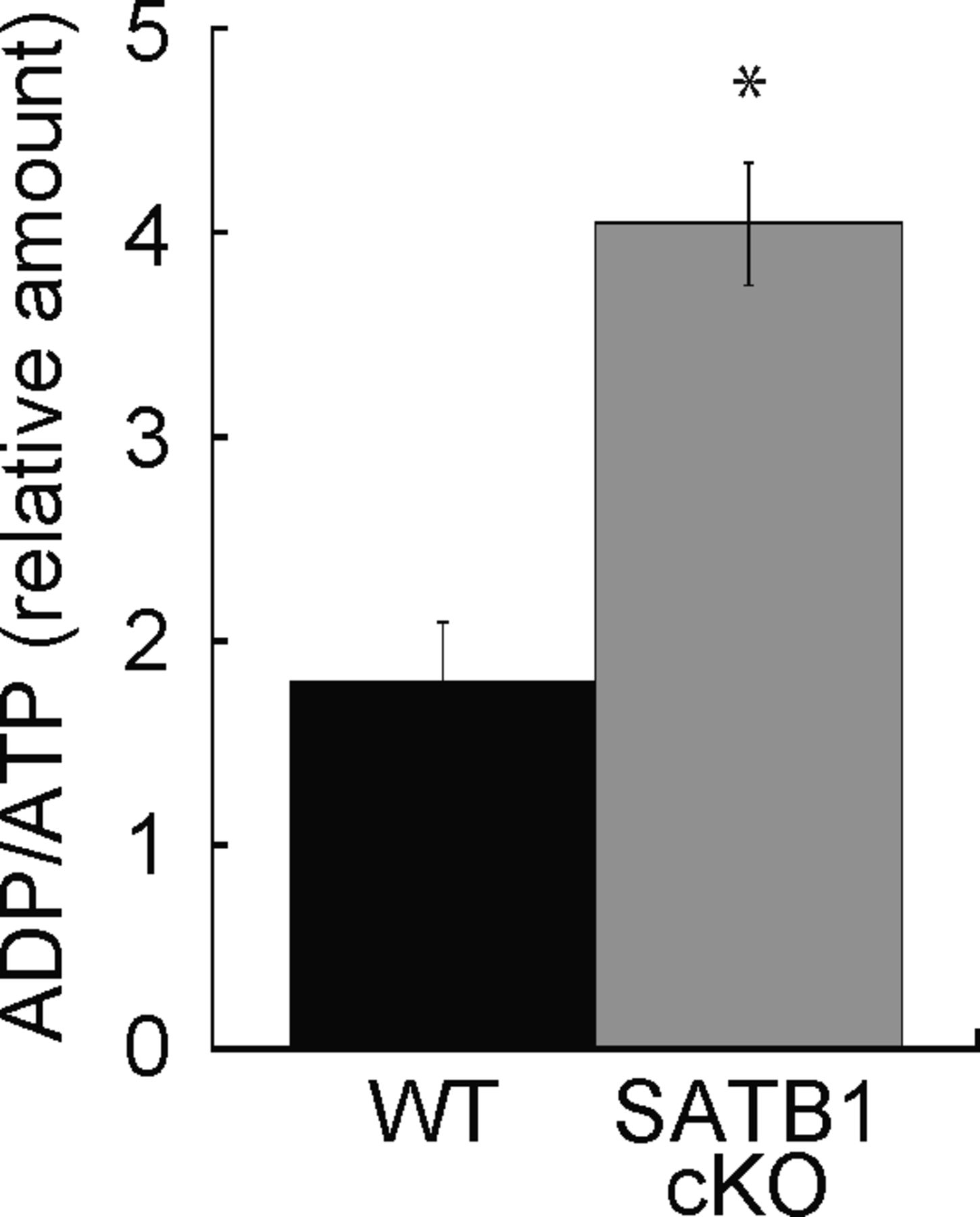
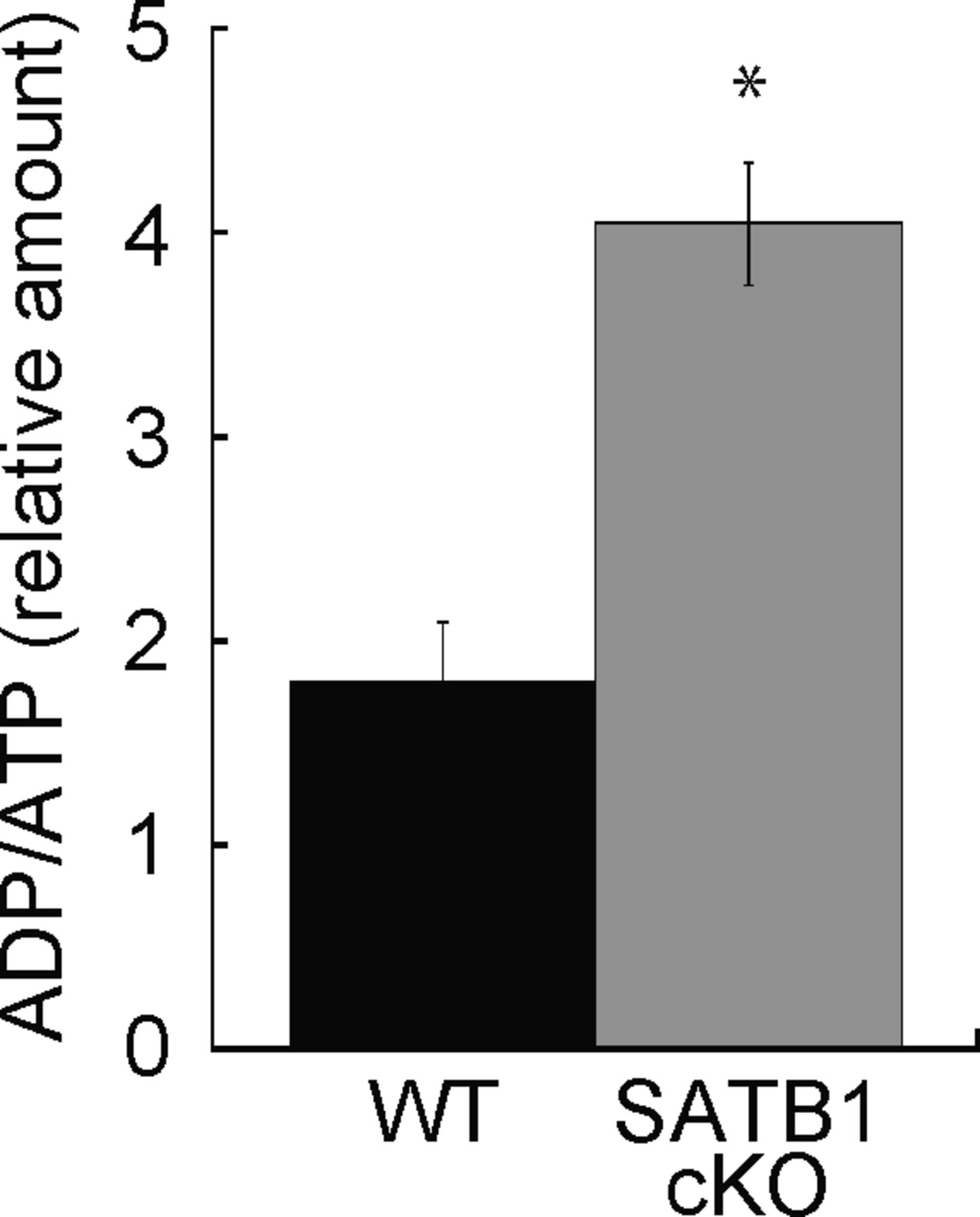
- Figure S3. ADP/ATP ratio in T cells.
Cellular ATP was assessed. Cell lysates were incubated with ADP assay buffer for 1 min and luminescence was measured (Lu-ADP). ADP/ATP ratio was calculated as Lu-ADP/Lu-ATP. Dye absorbance was measured in a plate reader at 450 nm. *P < 0.01 versus WT. N = 5. Data are shown as the means ± SD.
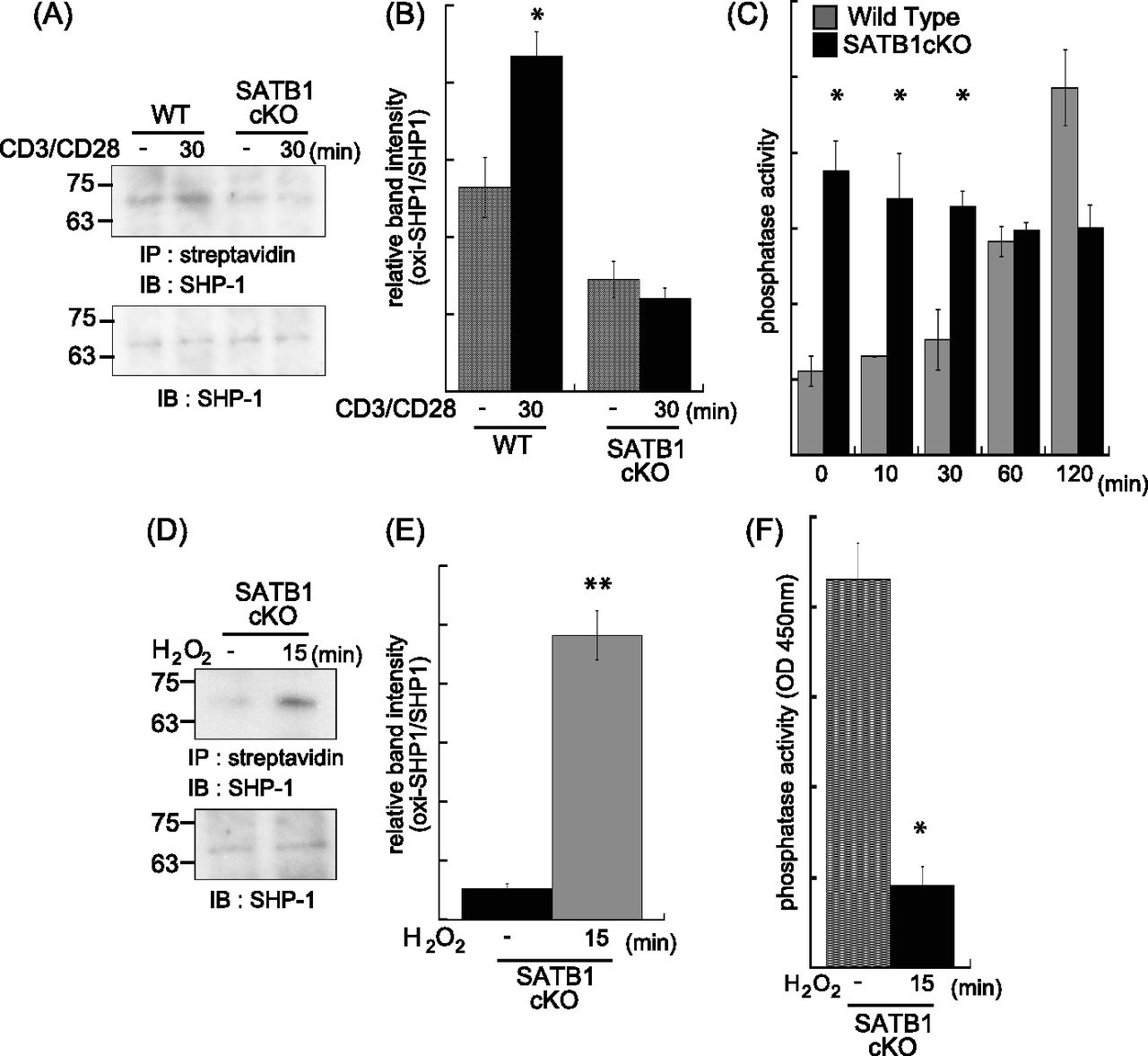
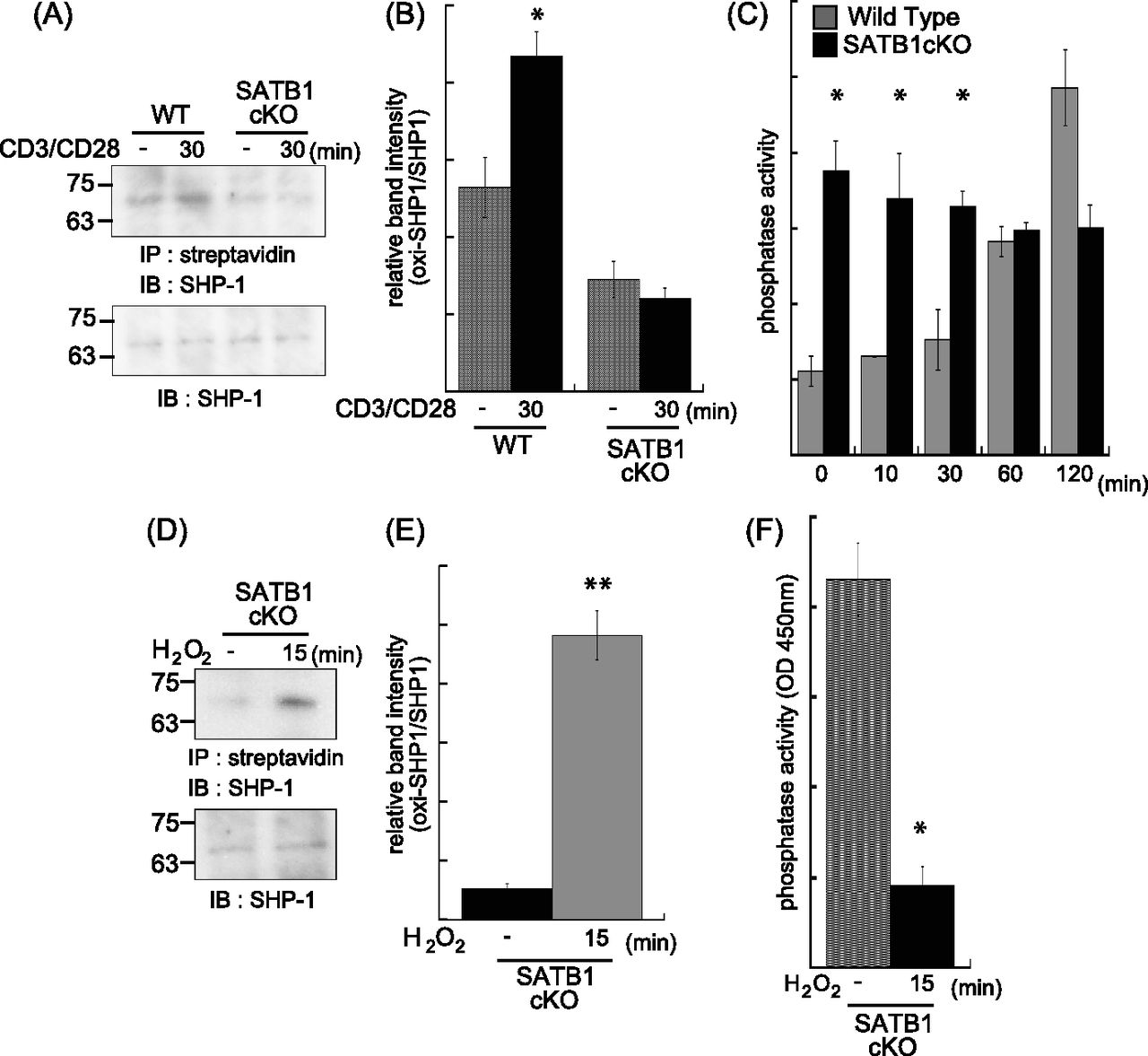
- Figure 3. Phosphatase SHP-1 is regulated by mtROS.
WT and SATB1cKO naïve CD4 T cells were stimulated with anti-CD3 and anti-CD28 antibodies for the indicated times and lysed in sample buffer. Oxidized proteins in the cell lysates were labeled with iodoaceto-PEG biotin and immunoprecipitated. (A) Blots were probed for anti–SHP-1 antibody. (B) Band intensities were quantified with ImageJ v. 1.49. (C) Naïve CD4 T Cells were incubated for 0, 10, 20, 30, 60, and 120 min with anti-CD3 and anti-CD28 antibodies and lysed in phosphatase assay buffer. Phosphatase activity was assayed by incubation with PY containing peptide and liberated phosphate was measured by malachite green absorption at 450 nm. (D, E) Lysates of SATB1-deficient naïve CD4 T cells were incubated in the absence or presence of H2O2 for 15 min. Oxidized SHP-1 was detected by immunoblotting with antibody against SHP-1 (D). (E) Band intensities were quantified with ImageJ v. 1.49 (E). (B, F) Phosphatase activity in the presence or absence of H2O2 was measured in (B). Data are shown as the means of three experiments performed in duplicate. *P < 0.01 versus WT. **P < 0.05 versus WT. N = 5. Data are shown as the means ± SD.
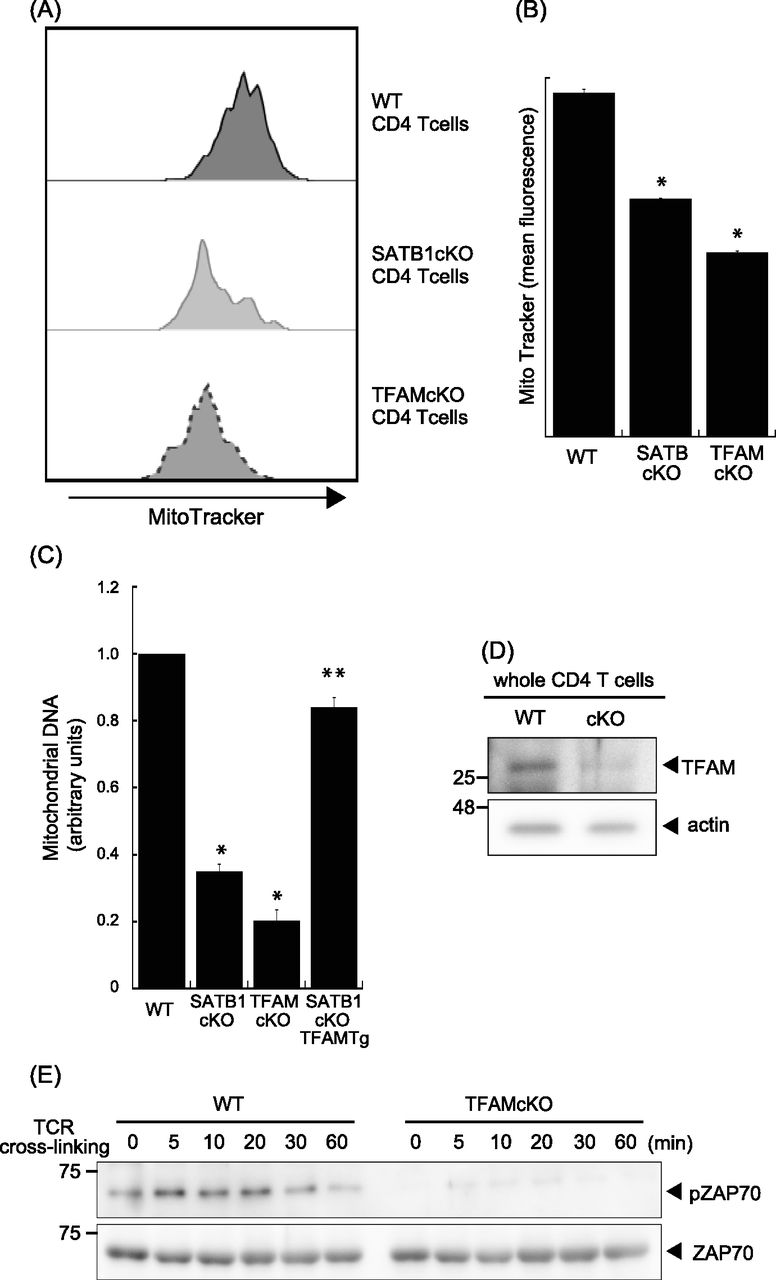
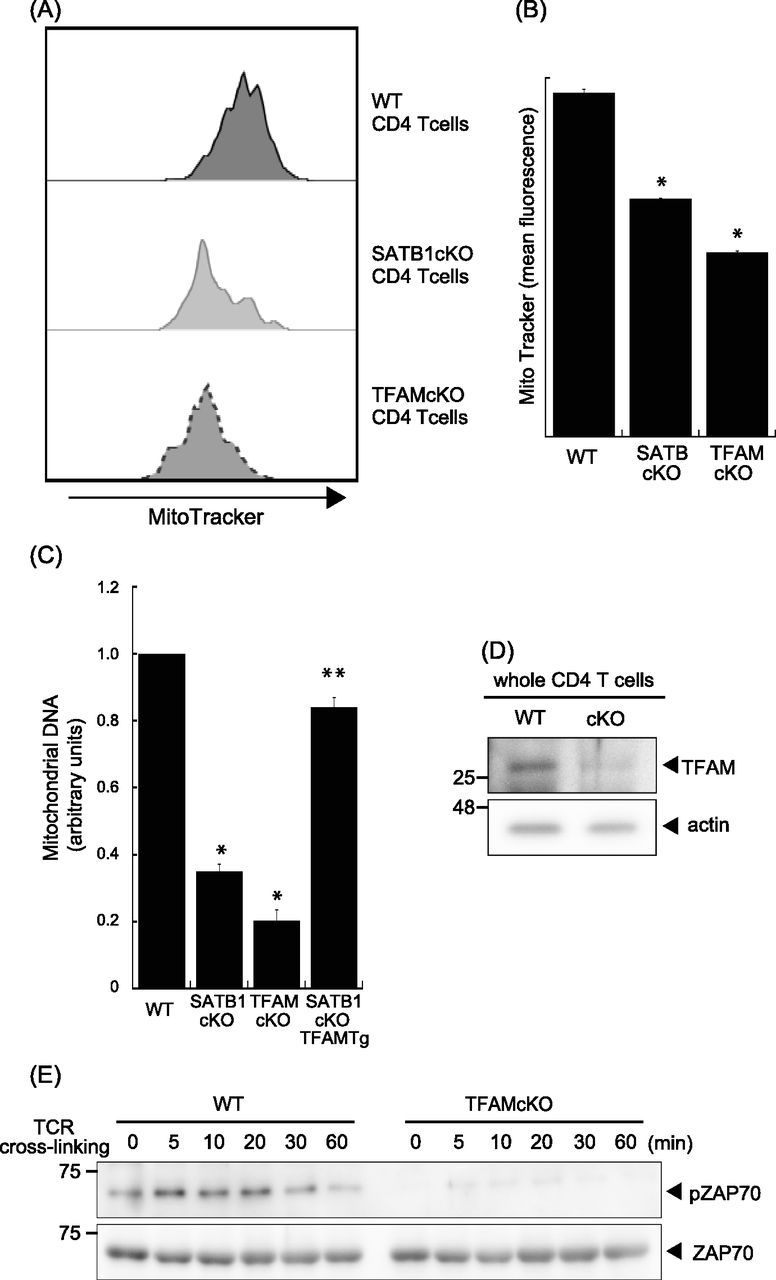
- Figure 4. Loss of TFAM expression reduces T-cell activity.
(A) Representative MitoTracker staining histograms. WT and SATB1cKO T cells were stained with MitoTracker and analyzed by FACS. (B) Quantification of the mean fluorescence of MitoTracker is shown. (C) Mitochondrial DNA copy numbers in CD4 T cells from WT, SATB1 cKO, TFAMcKO, and SATB1cKO-TFAMTg mice were measured by RT-qPCR. The mitochondrial DNA level was normalized to the nuclear DNA level in each sample. The normalized index is indicated as an arbitrary unit (WT = 1). (D) TFAM and β-actin levels in CD4 T-cell lysates were analyzed by immunoblotting using the respective antibodies. (E) CD4 T cells were prepared from WT and TFAMcKO mice, stimulated for the indicated times with anti-CD3 and anti-CD28 antibodies, lysed, analyzed by SDS–PAGE, and immunoblotted with antibodies against phospho-ZAP70, and ZAP70. Similar results were obtained for three independent experiments. *P < 0.01 versus WT. **P < 0.01 versus SATB1cKO T cells. N = 5. Data are shown as the means ± SD.
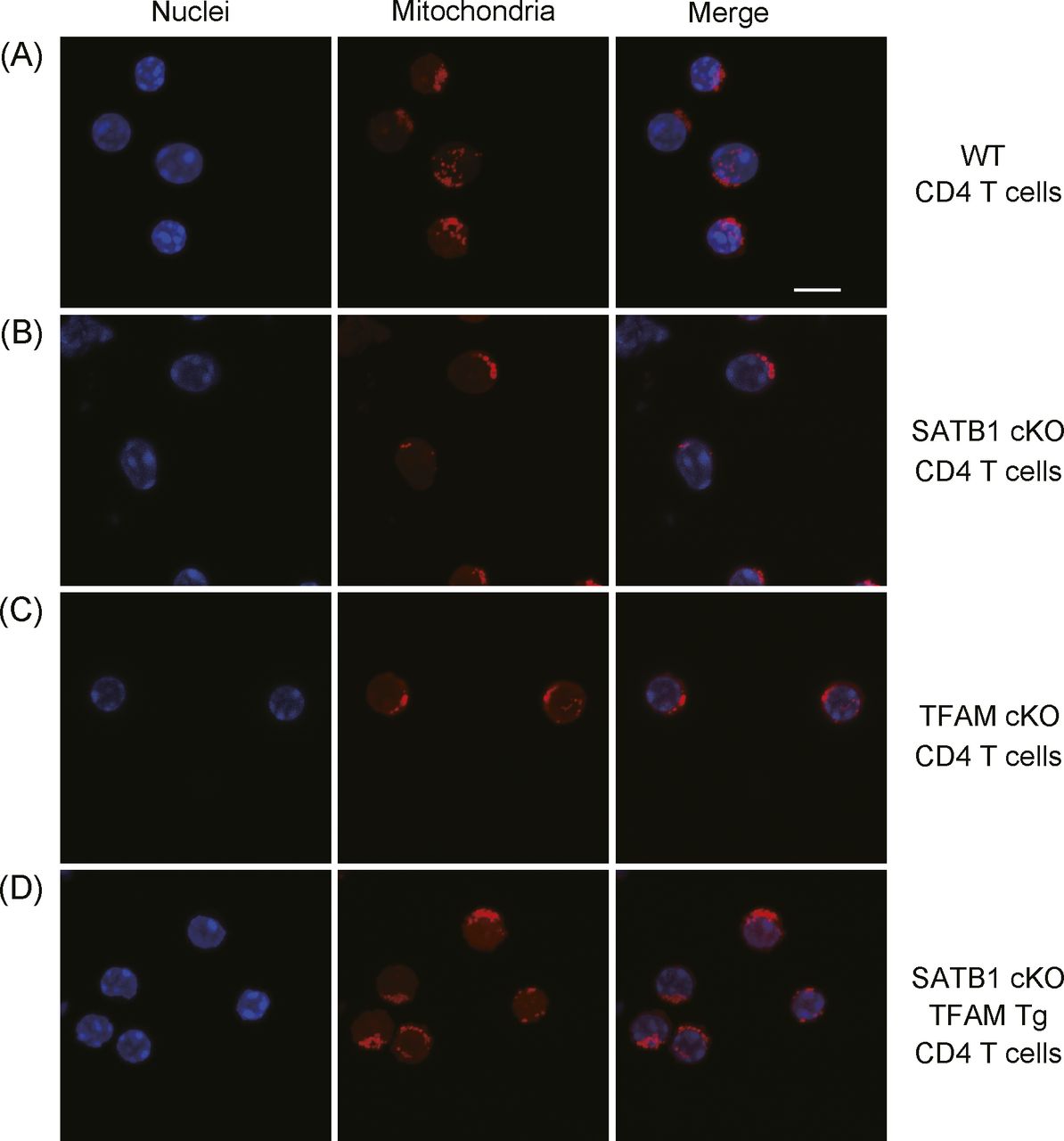
- Figure S4. Confocal imaging of mitochondria.
(A, B, C, D) WT CD4 T cells (A), SATB1cKO CD4 T cells (B), TFAMcKO CD4 T cells (C), and SATB1cKO-TFAMTg CD4 T cells (D) were stained with MitoTracker (mitochondria) and TO-PRO3 dye (nuclei) and analyzed by confocal microscopy. Scale bar, 10 μm. Data represent three independent experiments.
- Figure S5. TFAM mRNA expression was analyzed by RT-qPCR.
TFAM mRNA transcription levels were analyzed relative to β-actin. Data are shown as the means ± SD for three experiments. *P < 0.01 versus WT.
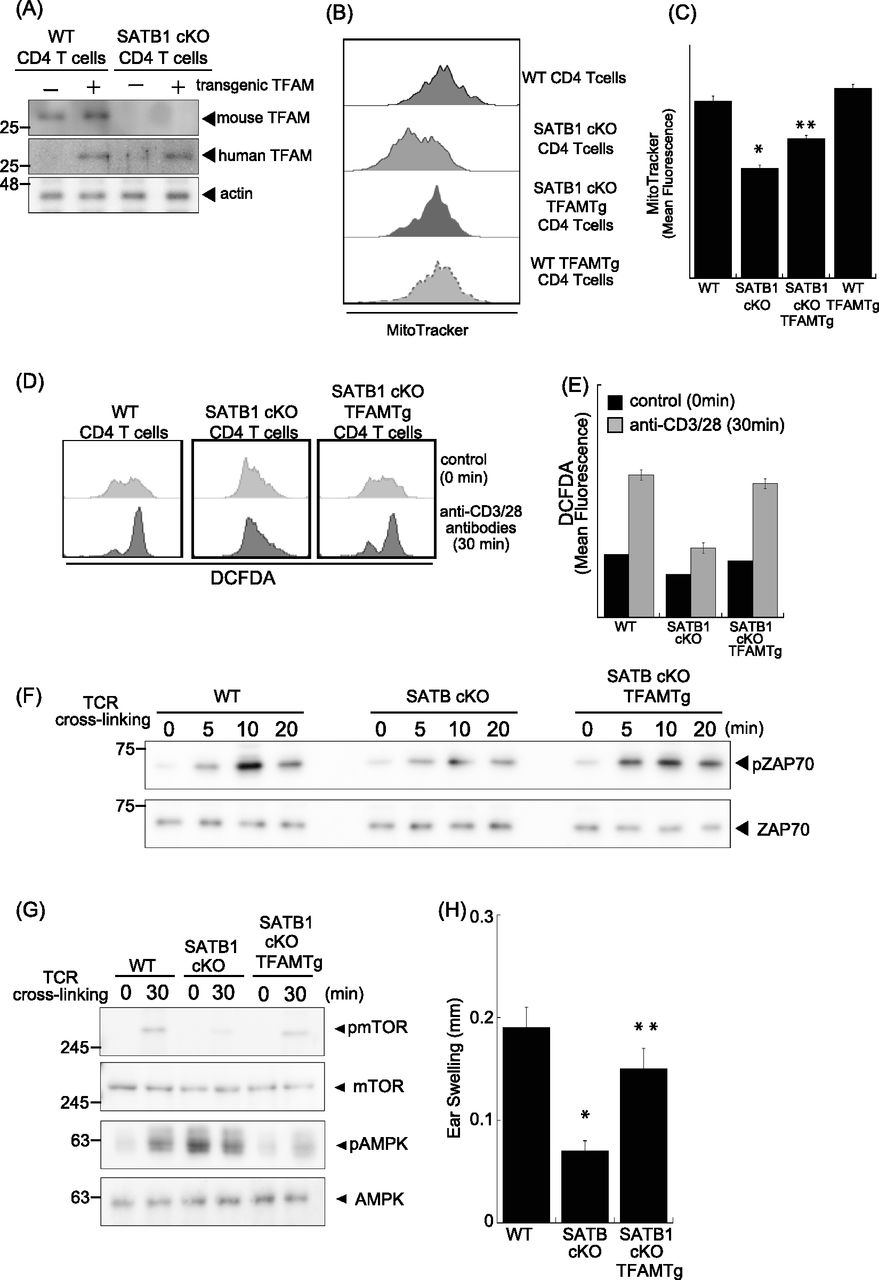
- Figure 5. TFAM expression rescues T-cell activity in SATB1cKO mice.
(A) Immunoblot analysis of transgenic TFAM and endogenous TFAM in SATB1-sufficient or SATB1-deficinet CD4 T cells. (B) Representative MitoTracker staining histograms. WT, SATB1cKO, SATB1cKO-TFAMTg, and WT-TFAMTg T cells were stained with MitoTracker and analyzed by FACS. (C) Quantification of the mean fluorescence of MitoTracker. *P < 0.01 versus WT. **P < 0.01 versus SATB1cKO T cells. N = 5. Data are shown as the means ± SD. (D) ROS levels in CD4 T cells from WT, SATB1cKO, and SATB1cKO-TFAMTg mice in control and TCR activation treatments were measured by DCFDA staining and flow cytometry. (E) Quantification of the mean fluorescence of DCFDA. (F, G) CD4 T cells were prepared from WT, SATB1cKO, and SATB1cKO-TFAMTg mice and functionally analyzed. T cells were stimulated with anti-CD3 and anti-CD28 antibodies for the indicated times, lysed, analyzed by SDS–PAGE, and immunoblotted with antibodies against phospho-ZAP70, ZAP70, phospho-mammalian target of rapamycin, phospho-AMPK, and AMPK. Similar results were obtained for three independent experiments. (H) Mice were sensitized with 7% PCI in acetone/olive oil. After 7 d, 1% (vol/vol) PCI was applied to the right ear and vehicle alone was applied to the left ear. Ear thickness was measured after 24 h. Means ± SD for 4–5 mice are indicated. *P < 0.01 versus WT. **P < 0.01 versus SATB1cKO T cells. N = 5. Data are shown as the means ± SD.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- No citing articles found.