



Article Figures & Data
Figures
- Figure S1. Identification of WWP1 as the prime E3 ligase which mono-ubiquitinates AMOTL2 at Lys-347/408.
(A) 293T cells were transfected with the indicated DNAs, then subjected to in vivo ubiquitination assays. Shown are the E3 ligases which were successfully expressed, and demonstrated a positive ubiquitination effect from the initial 160 E3 ligases from the ORF library (except for WWP2—a closely related family member of WWP1—which did not demonstrate a positive ubiquitination effect, and was included to serve as a negative control). (B) 293T cells were transfected with the indicated DNAs, then subjected to in vivo ubiquitination assays. (A) Shown are the E3 ligases which exhibited the greatest ubiquitination effect from (A). AMOTL2 2KR mutant was also included in this set, to assess whether the E3 ligase candidates cannot ubiquitinate this 2KR mutant. As can be seen, only WWP1 met this criteria.
- Figure 1. AMOTL2 is a mono-ubiquitination substrate of the E3 ligase WWP1.
(A) 293T cells were transfected with the indicated DNAs, then subjected to in vivo ubiquitination assays. (B) 293T cells were transfected with the indicated DNAs, then subjected to in vivo ubiquitination assays. (C) GST-WWP1 and AMOTL-V5 proteins purified from baculovirus were mixed with GST-UBE1, UbcH5c, and His-Ub as indicated and subjected to in vitro ubiquitination assay. M (molecular weight marker lane), CBB (Coomassie Brilliant Blue).
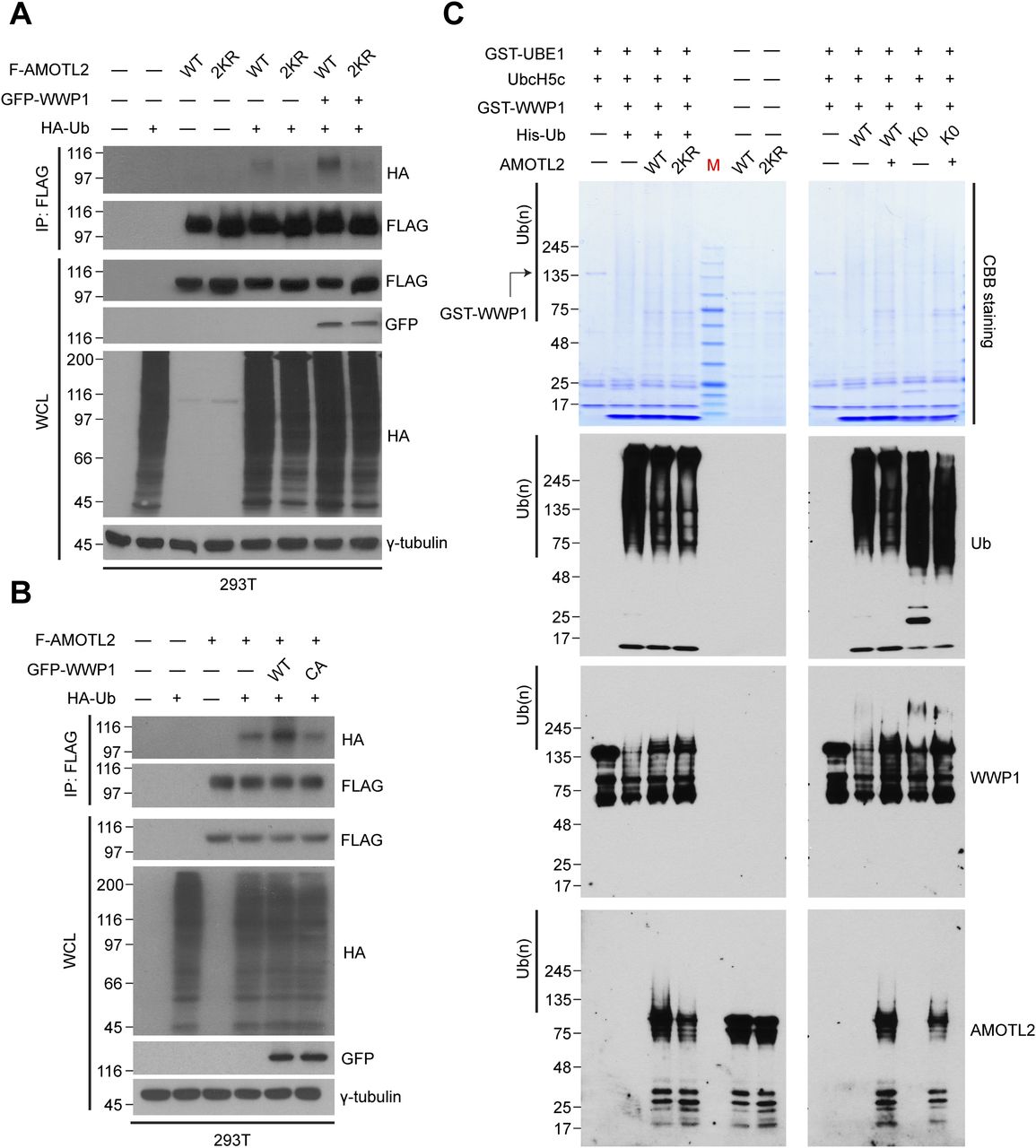
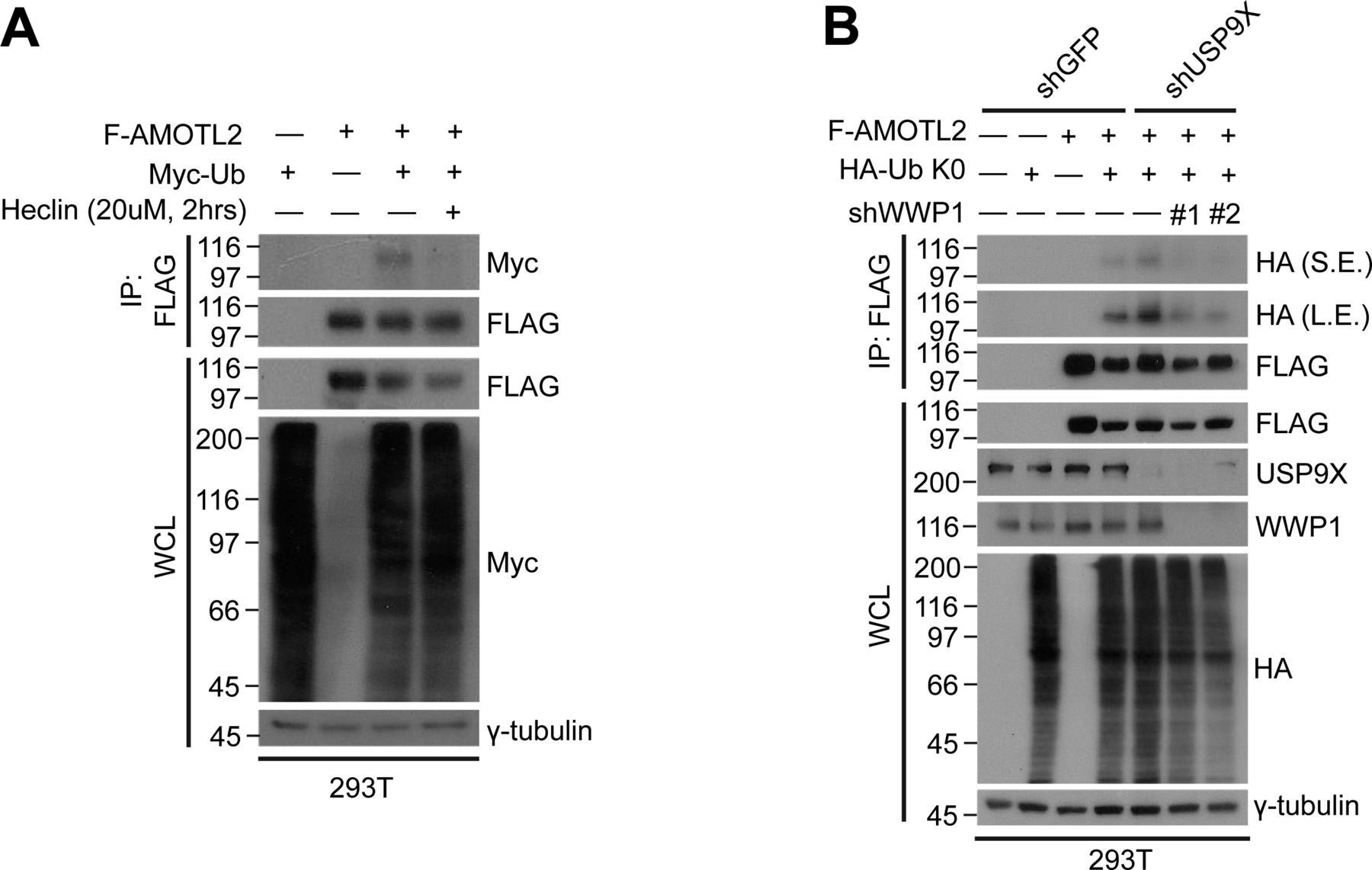
- Figure S2. AMOTL2 mono-ubiquitination is reversed upon heclin treatment or depletion of WWP1 in USP9X-deficient cells.
(A) 293T cells were transfected with the indicated combination of DNAs, and treated with either DMSO or 10 μM heclin for 2 h, as indicated. Extracts were then subjected to in vivo ubiquitination assays. (B) 293T cells stably expressing shRNAs against USP9X, WWP1, or a combination thereof were transfected with the indicated plasmids, and then the extracts were subjected to in vivo ubiquitination assays.
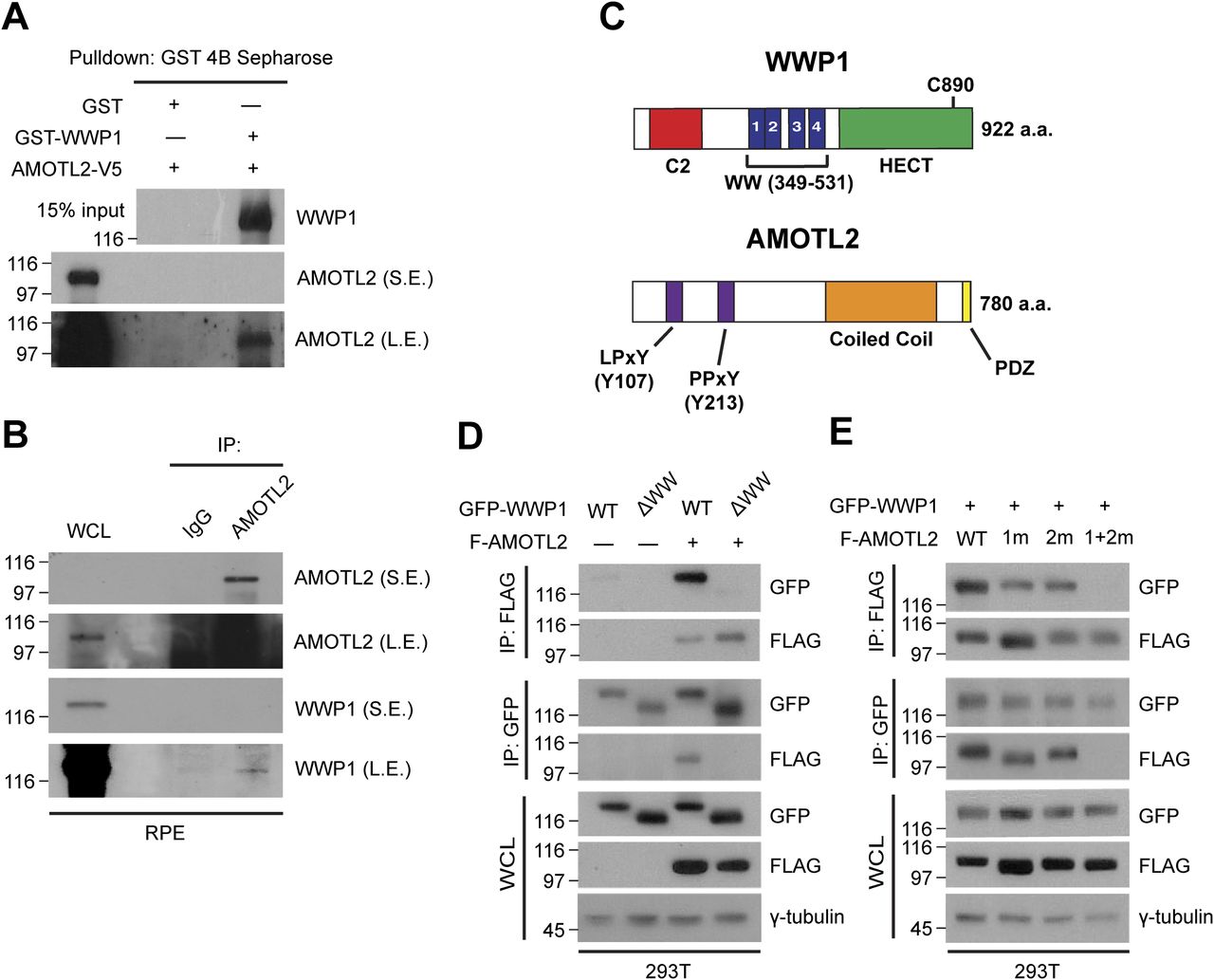
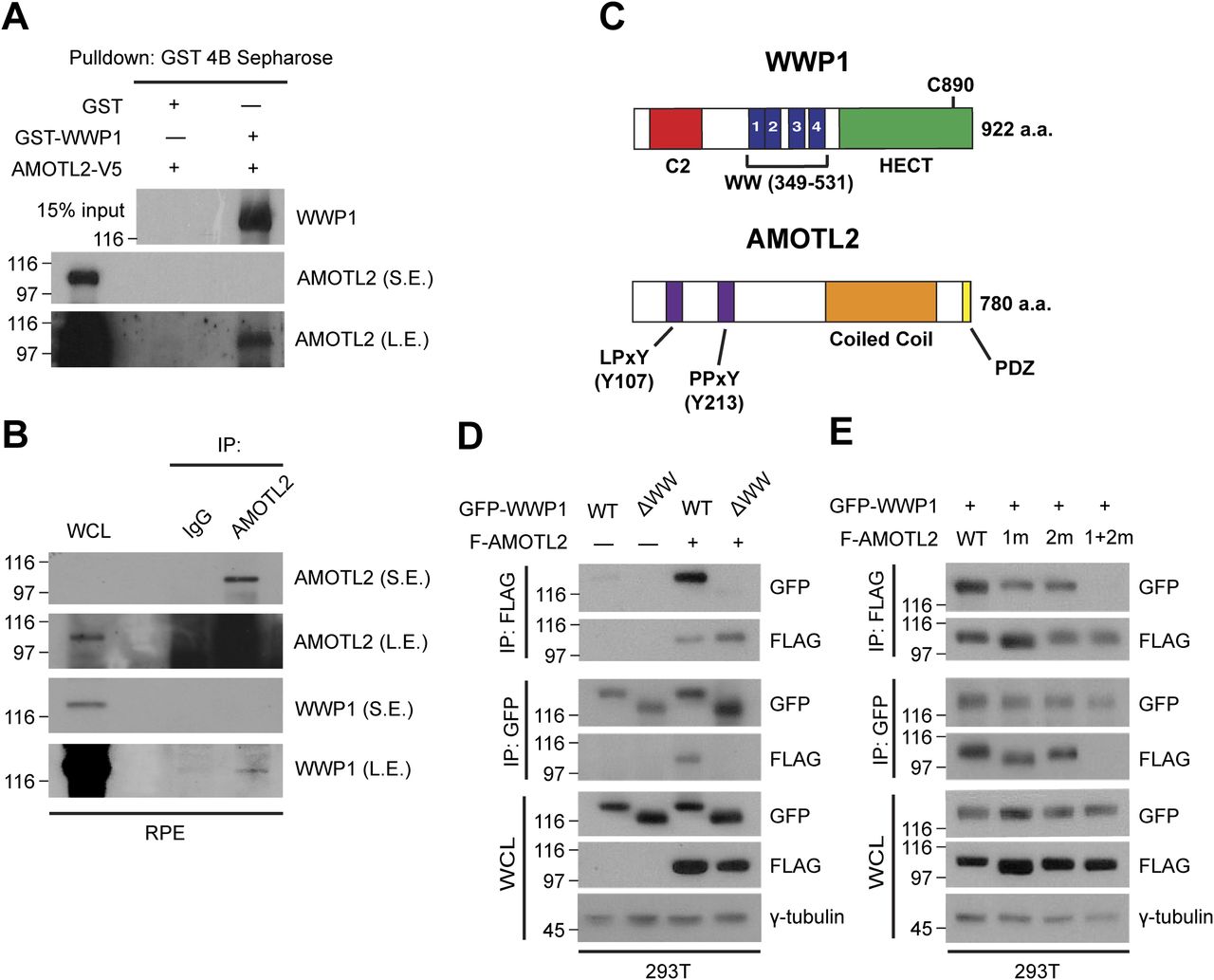
- Figure 2. WWP1 physically interacts with AMOTL2.
(A) GST-WWP1 and AMOTL-V5 proteins purified from baculovirus were subjected to in vitro binding assays using GST-4B Sepharose to pull down GST or GST-WWP1. S.E., short exposure; L.E., long exposure. (B) Confluent RPE-1 cells were lysed and endogenous proteins were co-immunoprecipitated with an anti-AMOTL2 antibody. (C) Schematic diagram illustrating the functional domains of WWP1 and AMOTL2. The amino acids shown in brackets represent sites either deleted or point-mutated in WWP1 and AMOTL2, respectively. (D) 293T cells were transfected with the indicated DNAs then subjected to reciprocal co-immunoprecipitation assays. (E) 293T cells were transfected with the indicated DNAs then subjected to reciprocal co-immunoprecipitation assays.
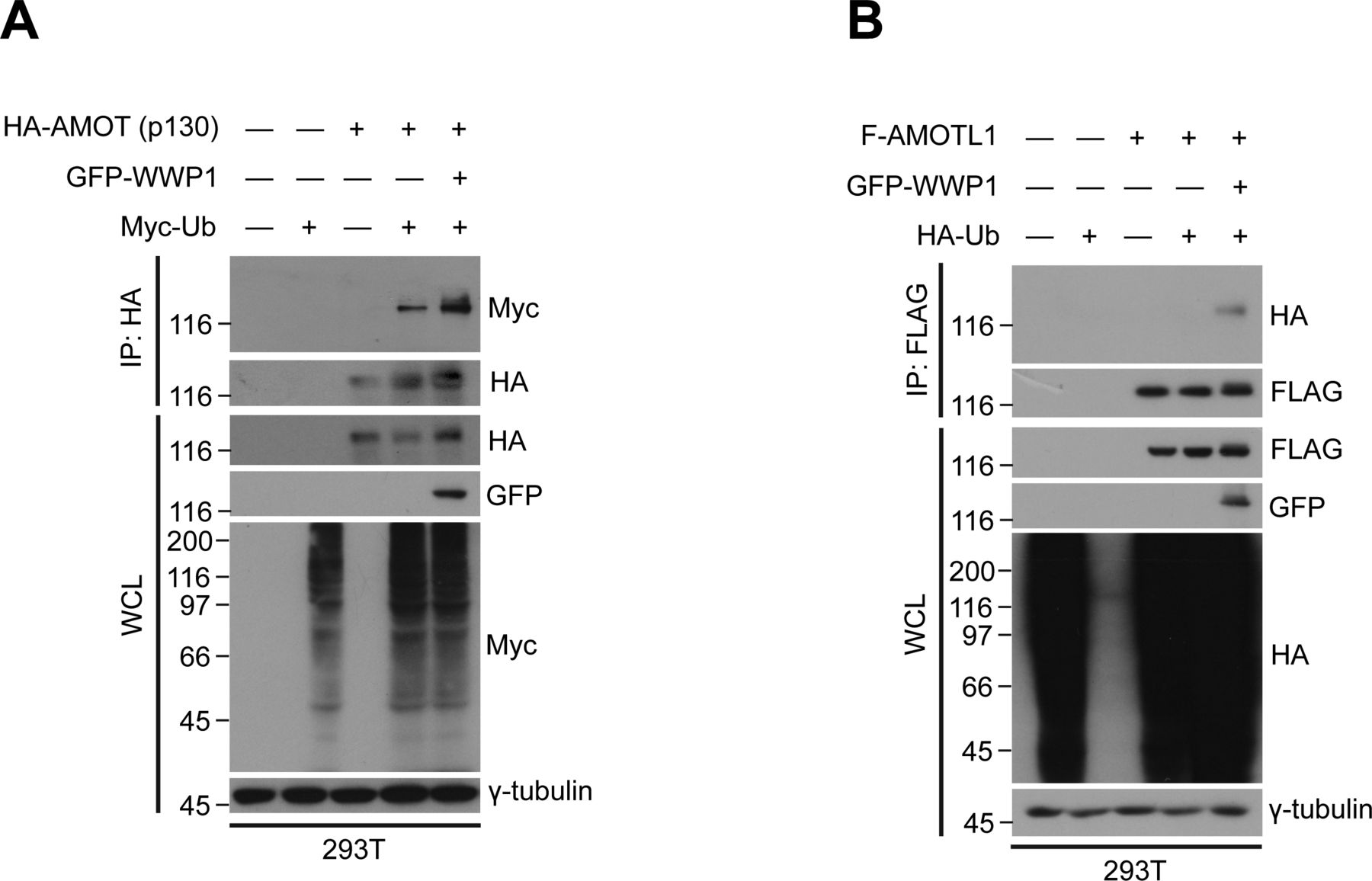
- Figure S3. AMOTL2 isoforms are mono-ubiquitinated by WWP1.
(A) 293T cells were transfected with GFP-WWP1 and Myc-Ub together with HA-AMOT (p130), then subjected to in vivo ubiquitination assays followed by Western blot analysis. (B) 293T cells were transfected with GFP-WWP1 and HA-Ub together with FLAG-AMOTL1, then subjected to in vivo ubiquitination assays followed by Western blot analysis.
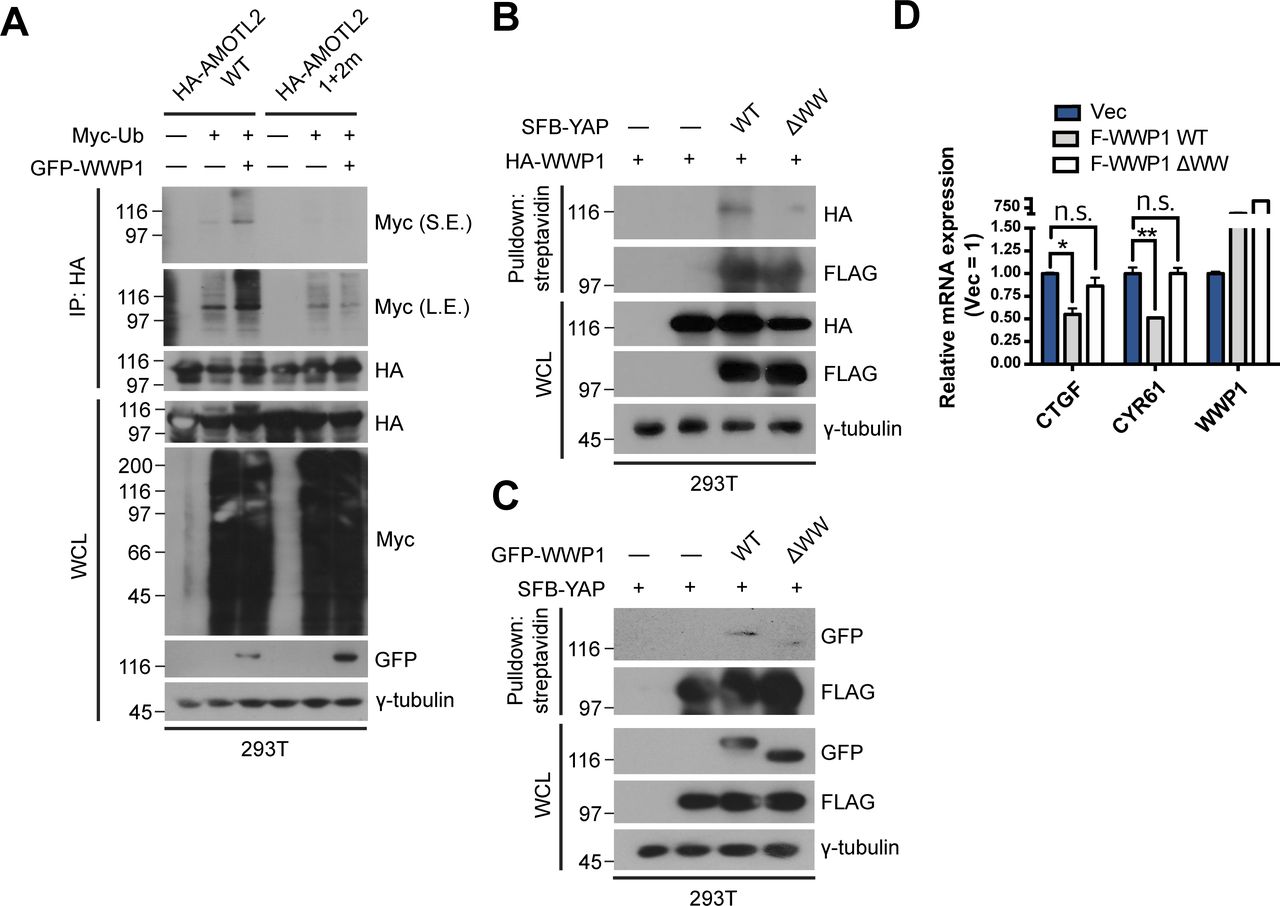
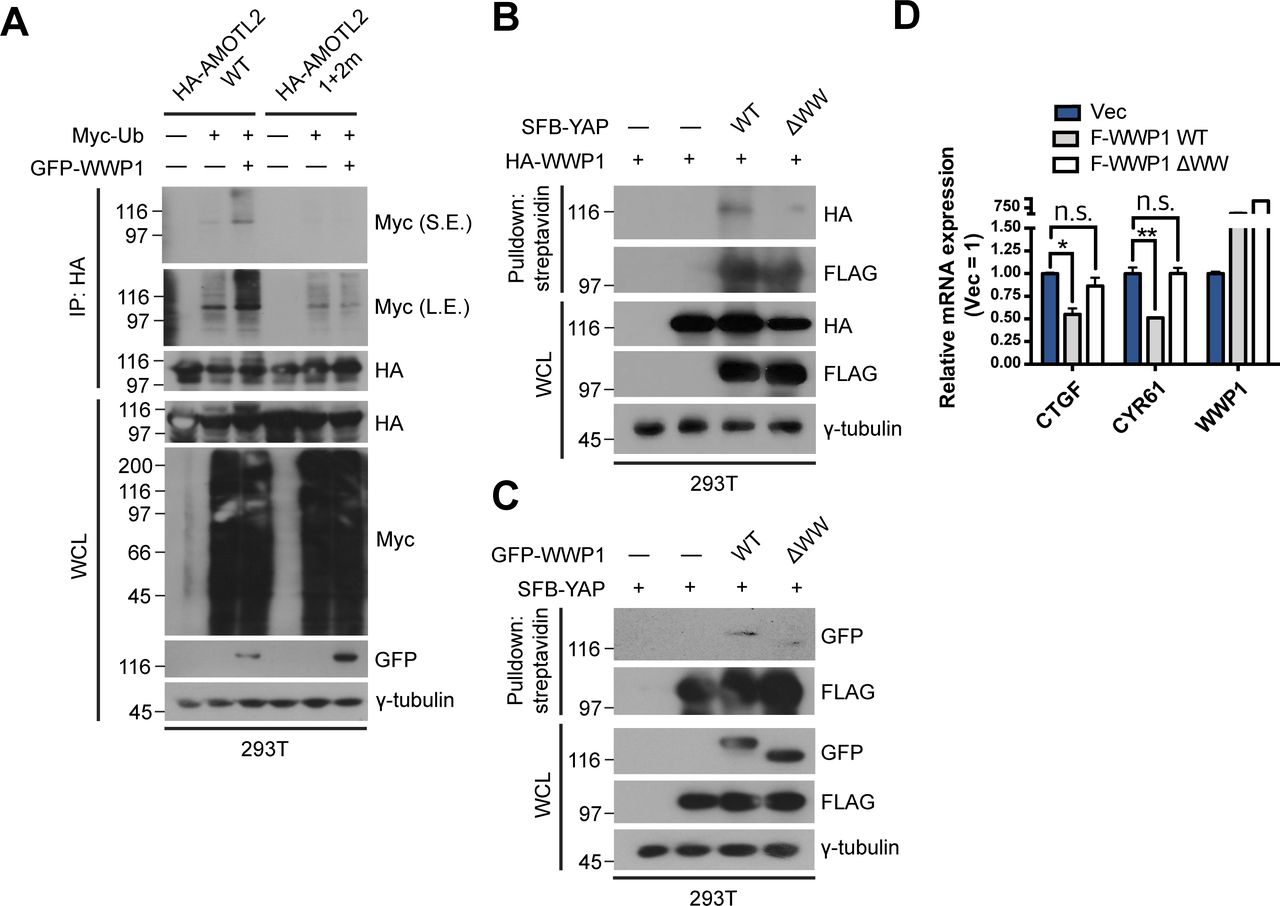
- Figure S4. Binding of WWP1 is important for AMOTL2 mono-ubiquitination and ultimately YAP inactivation.
(A) 293T cells were transfected with the indicated combination of DNAs, and extracts were then subjected to in vivo ubiquitination assays. (B) 293T were cells transfected with the indicated DNAs, then proteins in the resulting lysates were pulled down using streptavidin beads followed by Western blot analysis. (C) 293T were cells transfected with the indicated DNAs, then proteins in the resulting lysates were pulled down using streptavidin beads followed by Western blot analysis. (D) MCF10A cells stably expressing FLAG-WWP1 WT or ΔWW were cultured under sparse conditions; mRNA extracts were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; *P < 0.05, **P < 0.01, n.s. not significant; unpaired t test).
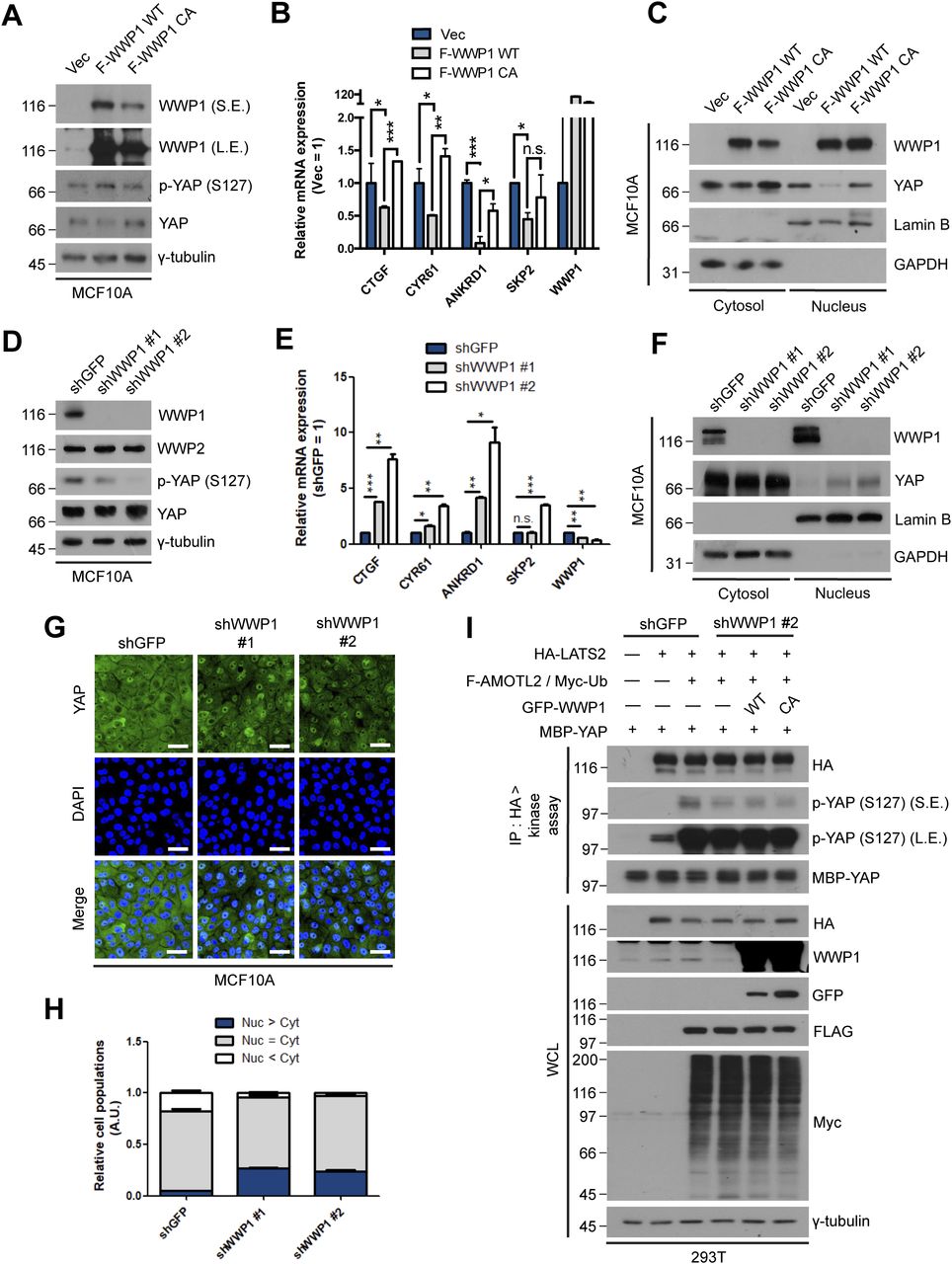
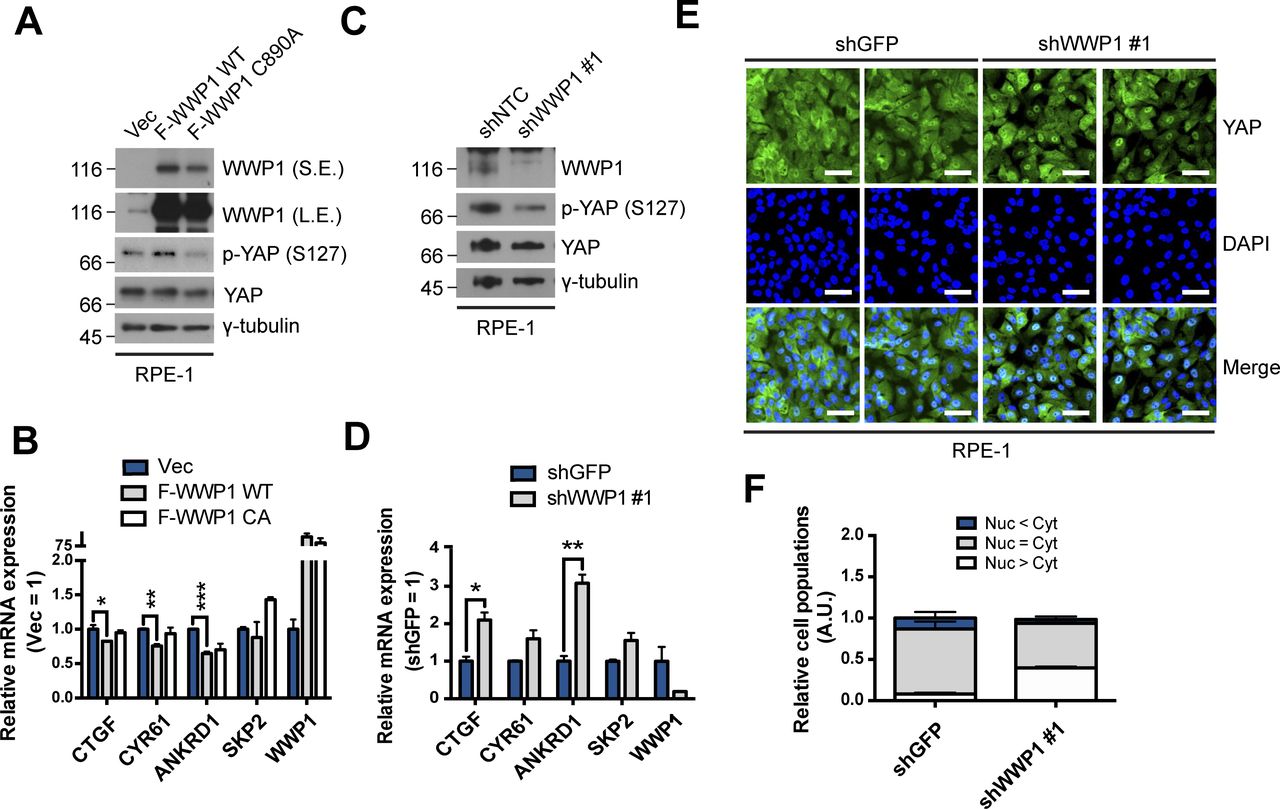
- Figure 3. WWP1 restrains YAP nuclear localization and suppresses its transcriptional activity.
(A) MCF10A cells were stably transduced with control, FLAG-WWP1 WT, or C890A mutant retroviruses, re-seeded at sparse conditions, then analyzed for the indicated proteins by Western blotting. (A, B) mRNAs extracted from cells prepared as in (A) were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; *P < 0.05, **P < 0.01, ***P < 0.001; unpaired t test). (A, C) Cells prepared as in (A) were fractionated into cytosolic and nuclear extracts, and analyzed by Western blotting for the indicated proteins. GAPDH and lamin B serve as controls for the cytosolic and nuclear fractions, respectively. (D) MCF10A cells stably transduced with shGFP (control) or shWWP1 were seeded under confluent conditions,, then lysed and analyzed for the indicated proteins by Western blotting. (D, E) mRNAs from cells prepared as in (D) were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; *P < 0.05, ***P < 0.001; unpaired t test). (D, F). Cells prepared as in (D) were fractionated into cytosolic and nuclear extracts, and analyzed by for the indicated proteins by Western blotting. GAPDH and lamin B serve as controls for the cytosolic and nuclear fractions, respectively. (D, G). Representative images of anti-YAP immunofluorescence in cells prepared as in (D). Green, YAP; blue, DAPI. Scale bar, 50 μm. (G, H) Quantification of relative cell populations shown in (G). At least four independent areas were counted. (I) 293T cells stably transduced with shGFP (control) or shWWP1 were transfected with the indicated DNAs, then extracts were subjected to LATS2 in vivo kinase assays using bacterially purified maltose-binding protein-YAP as a substrate.
- Figure S5. WWP1 restrains YAP nuclear localization and suppresses its transcriptional activity in RPE-1 cells.
(A) RPE-1 cells were stably transduced with control, FLAG-WWP1 WT, or C890A mutant retroviruses, re-seeded at sparse conditions, then analyzed for the indicated proteins by Western blotting. (A, B) mRNAs extracted from cells prepared as in (A) were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; *P < 0.05, **P < 0.01, ***P < 0.001; unpaired t test). (C) RPE-1 cells were stably transduced with shControl (targeting GFP) or shWWP1 lentiviruses, re-seeded at dense conditions, and then analyzed for the indicated proteins by Western blotting. (C, D) mRNAs extracted from cells prepared as in (C) were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; *P < 0.05, ***P < 0.001; unpaired t test). (C, E) Representative images of anti-YAP immunofluorescence in cells prepared as in (C). Green, YAP; blue, DAPI. Scale bar, 50 μm. (E, F) Quantification of relative cell populations shown in (E). At least four independent areas were counted.
- Figure S6. WWP1 suppresses cell proliferation.
(A) MCF10A cells stably transduced with shGFP (control) or shWWP1 lentiviruses and shRNA-resistant WT or C890A WWP1 retroviruses were seeded to confluence, then BrdU (5 μg/ml) was incorporated for 1 h. Representative images of anti-BrdU immunofluorescence is shown. Green, BrdU; blue, DAPI. Scale bar, 50 μm. (A, B) Quantification of relative cell populations shown in (A). At least four independent areas were counted. Data are expressed as means ± SEM (error bars; *P < 0.05, ***P < 0.001, n.s. not significant; unpaired t test). (A, C) Western blot analysis of samples prepared as in (A), showing successful depletion and subsequent rescue of WWP1.
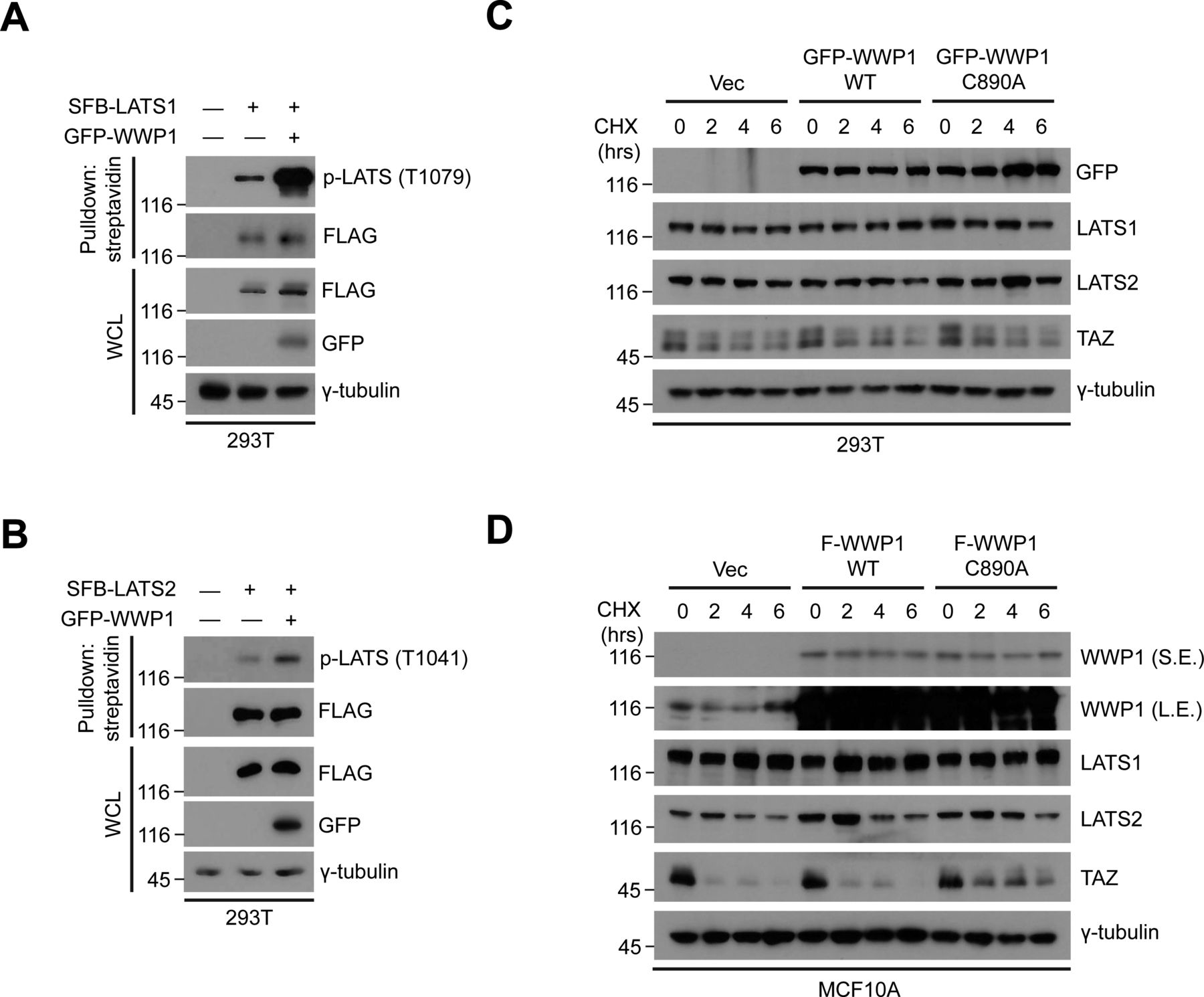
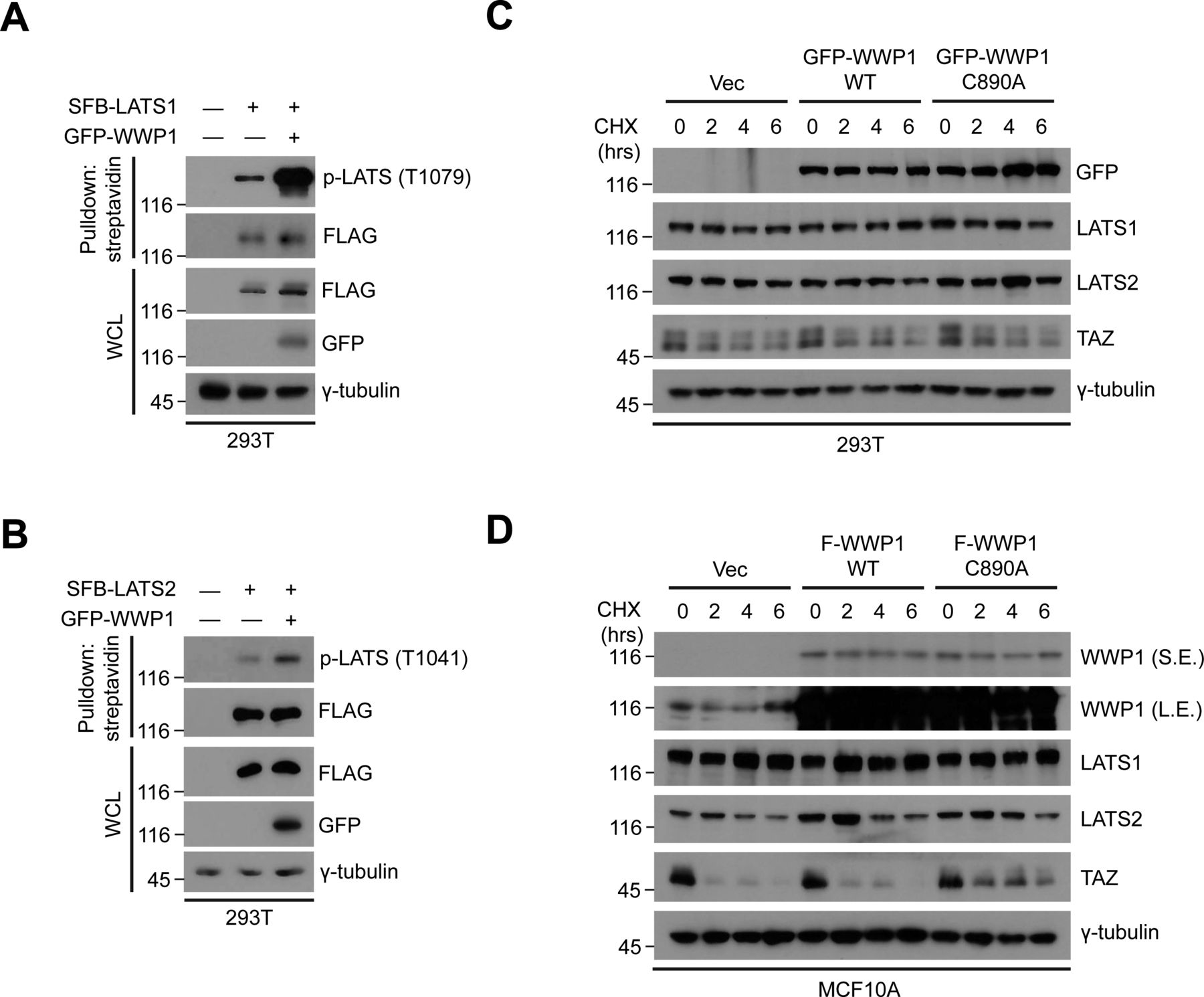
- Figure S7. WWP1 promotes the phosphorylation of LATS1/2 without affecting their stability.
(A) 293T were cells transfected with the indicated DNAs including SFB-LATS1, then proteins in the resulting lysates were pulled down using streptavidin beads followed by Western blot analysis to probe for phospho-LATS. (B) 293T were cells transfected with the indicated DNAs including SFB-LATS2, then proteins in the resulting lysates were pulled down using streptavidin beads followed by Western blot analysis to probe for phospho-LATS. (C) 293T cells transfected with GFP-WWP1 WT or C890A were subjected to cycloheximide (CHX) chase assay for the indicated times. The resulting extracts were analyzed by Western blotting. Immunoblot for TAZ, which has a relatively short half-life, was included to serve as a positive control for the experiment. (D) MCF10A cells stably transduced with FLAG-WWP1 WT or C890A were subjected to cycloheximide (CHX) chase assay for the indicated times. The resulting extracts were analyzed by Western blotting. Immunoblot for TAZ, which has a relatively short half-life, was included to serve as a positive control for the experiment.
- Figure 4. WWP1 promotes AMOTL2 phosphorylation by LATS.
(A) 293T cells stably transduced with shGFP (control) or shWWP1 lentiviruses were transfected with the indicated DNAs, then lysates were analyzed by Western blotting. (B) 293T cells were transfected with the indicated DNAs, then lysates were analyzed by Western blotting. (C) 293T cells were transfected with the indicated DNAs, then lysates were analyzed by Western blotting. (D) Control or CRISPR/Cas9-mediated LATS1/2-knockout 293T cells were transfected with the indicated DNAs, then lysates were subjected to in vivo ubiquitination assay.
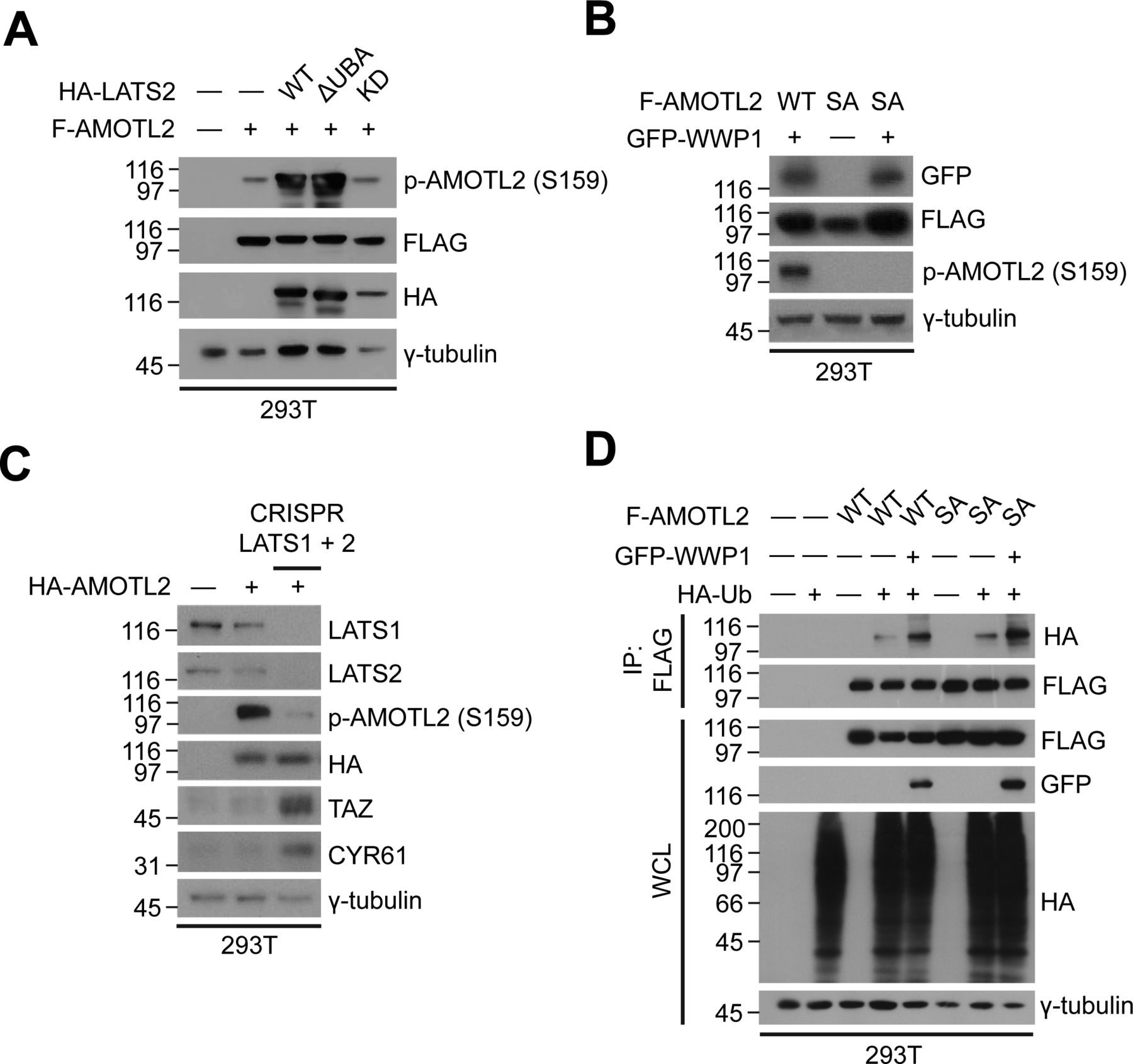
- Figure S8. Analysis of AMOTL2 phosphorylation.
(A) 293T cells were transfected with FLAG-AMOTL2 and HA-LATS WT, ΔUBA, or kinase dead, and extracts were analyzed for the indicated proteins by Western blotting. (B) 293T cells were transfected with GFP-WWP1 and WT or S159A FLAG-AMOTL2, and extracts were analyzed for the indicated proteins by Western blotting. (C) Wild-type or CRISPR/Cas9–mediated LATS1/2 double-knockout 293T cells were transfected with HA-AMOTL2, and extracts were analyzed for the indicated proteins by Western blotting. (D) 293T cells were transfected with WT or S159A FLAG-AMOTL2 together with GFP-WWP1 and HA-Ub, and extracts were subjected to in vivo ubiquitination assays followed by Western blot analysis.
- Figure 5. AMOTL2 mono-ubiquitination promotes LATS activation.
(A) 293T cells stably transduced with shGFP (control) or shWWP1 lentiviruses were transfected with the indicated DNAs, then lysates were pulled down using streptavidin agarose beads. SFB: tag comprising (in order) S-protein tag, FLAG tag, and SBP (streptavidin-binding peptide) tag. (B) 293T cells stably transduced with shGFP (control) or shWWP1 lentiviruses were transfected with the indicated DNAs, then lysates were immunoprecipitated with anti-FLAG antibody. #denotes endogenous WWP1 and *denotes exogenous GFP-WWP1. (C) 293T cells were transfected with the indicated DNAs, then extracts were immunoprecipitated with anti-FLAG antibody. (D) 293T cells were transfected with HA-LATS2, then extracts were subjected to LATS2 in vivo kinase assays using bacterially purified maltose-binding protein-YAP as substrate. Reactions were supplemented with 2 μg of baculovirus-purified free AMOTL2 or Ub-AMOTL2, obtained as a product of in vitro ubiquitination. (E) 293T cells were transfected with HA-tagged WT or ΔUBA LATS2 (Ub-acceptor domain deletion mutant), then extracts were subjected to in vivo kinase assays using bacterially purified maltose-binding protein-YAP as a substrate. (D) Reactions were supplemented with increasing molar amounts of Ub-AMOTL2 (0, 1, 2.5, and 6 μg), prepared as in (D).
- Figure S9. Isolation of mono-ubiquitinated AMOTL2.
GST-WWP1 and AMOTL-V5 proteins purified from baculovirus were mixed with GST-UBE1, UbcH5c, and His-Ub K0 as indicated and subjected to in vitro ubiquitination assay. However, in order to isolate AMOTL2 from WWP1, GST-WWP1 was initially bead-bound before reaction. Then after in vitro ubiquitination, the beads and supernatant samples were separately analyzed by SDS–PAGE. As can be seen, we could detect ubiquitinated AMOTL2 corresponding to its natural size, devoid of smearing or contamination from WWP1. Therefore, we used this supernatant fraction to supplement the LATS kinase assay shown in Fig 5D.
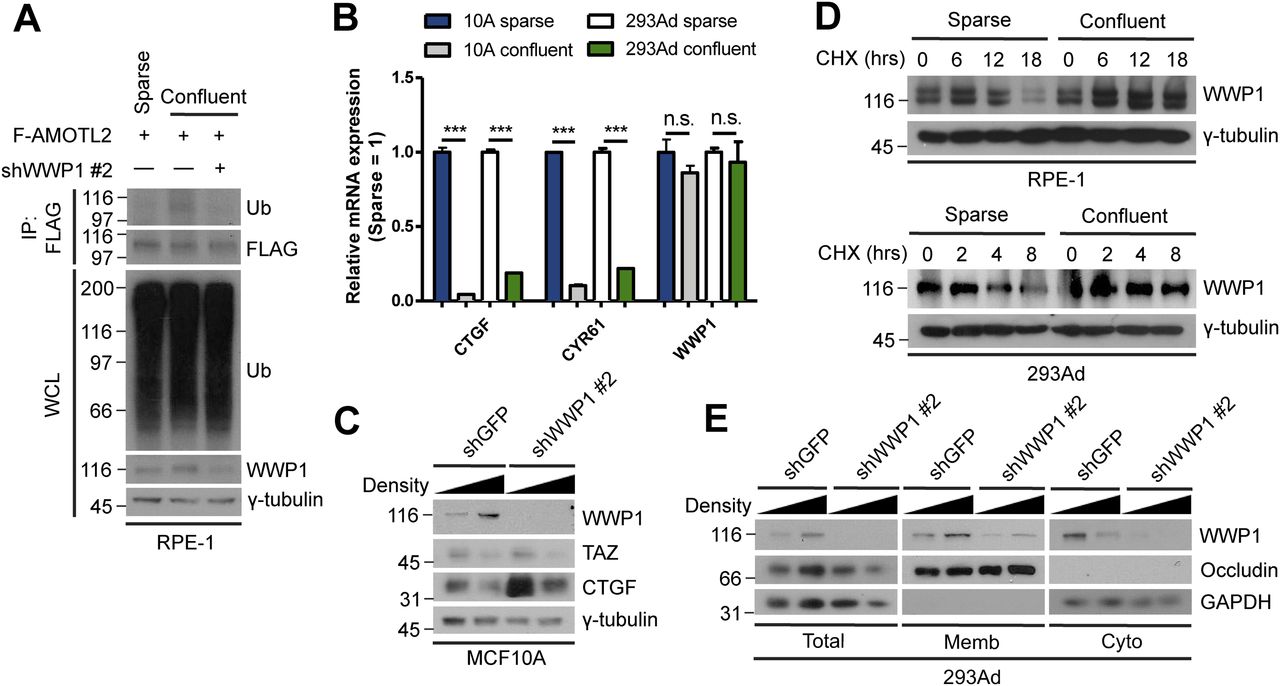
- Figure 6. High cell density promotes WWP1 stabilization and activation.
(A) RPE-1 cells were stably transduced with FLAG-AMOTL2 and either control or WWP1 shRNAs, re-seeded to a sparse or confluent density, then subjected to in vivo ubiquitination assays. (B) mRNAs isolated from MCF10A and 293Ad cells, seeded at either a sparse or confluent density, were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; ***P < 0.001, n.s. not significant; unpaired t test). (C) MCF10A cells stably transduced with shGFP (control) or shWWP1 lentiviruses were seeded at either a sparse or confluent density and extracts were analyzed by Western blotting. (D) RPE-1 and 293Ad cells were seeded at either a sparse or confluent density, then a cycloheximide (CHX) chase assay was performed for the indicated times. The resulting extracts were analyzed by Western blotting. (E) 293Ad cells were seeded at either sparse or confluent densities and extracts were subjected to S100/P100 membrane-cytosol fractionation assays. The resulting fractions were analyzed for the indicated proteins by Western blotting.
- Figure 7. WWP1 localizes to the Crumbs polarity complex via AMOTL2.
(A) MCF10A cells transfected with GFP-WWP1 WT or ΔWW were immunostained for GFP and PALS1, and serial z-stack images along the x-plane were overlaid by orthogonal projection. Green, GFP; red, PALS1; blue, Hoescht. Scale bar, 50 μm. (B) 293T cells were transfected with the indicated combination of DNAs, then subjected to SBP pull-down assays using streptavidin agarose beads followed by Western blot analysis. (C) 293T cells were transfected with the indicated combination of DNAs, then subjected to SBP pull-down assays using streptavidin agarose beads followed by Western blot analysis. (D) 293T cells were transfected with the indicated combination of DNAs, then subjected to SBP pull-down assays using streptavidin agarose beads followed by Western blot analysis. (E) 293T cells were stably transduced with either control or Crumbs3 shRNAs, then transfected with the indicated DNAs. The resulting extracts were subjected to in vivo ubiquitination assays, followed by Western blot analysis. (F) mRNAs isolated from MCF10A cells stably transduced with control or Crumbs3 shRNAs, then re-seeded at a sparse or confluent density were analyzed for the indicated genes by qRT-PCR, and expression levels were normalized to those of GAPDH mRNA (n = 4). Data are expressed as means ± SEM (error bars; *P < 0.05, ***P < 0.001, n.s. not significant; unpaired t test).
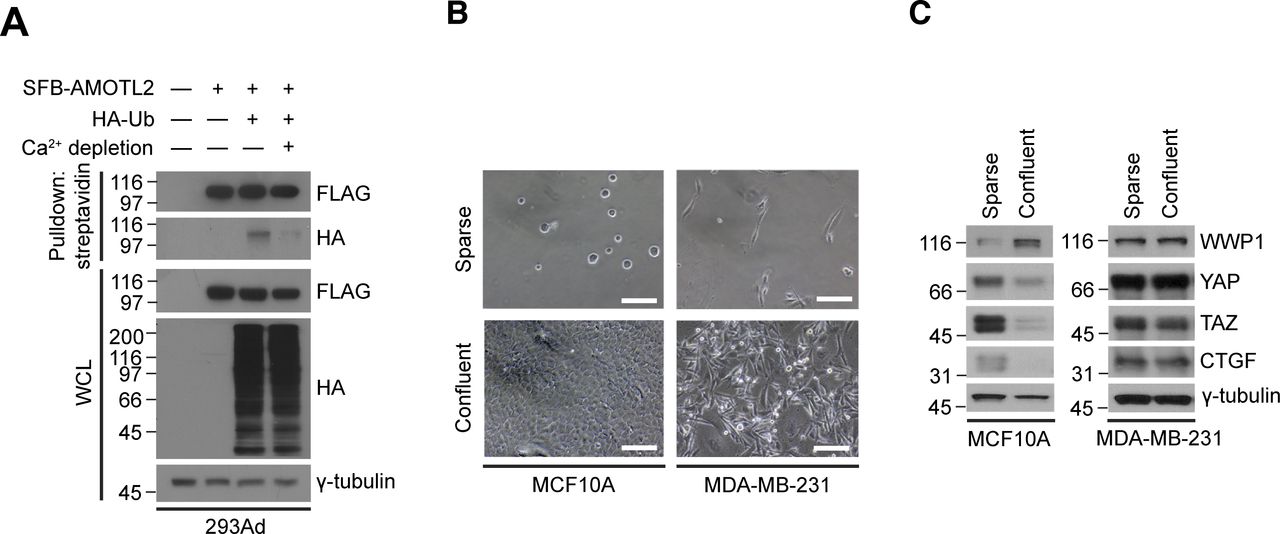
- Figure S10. Cell-cell contact is important for AMOTL2 mono-ubiquitination.
(A) 293Ad cells were transfected with the indicated DNAs, then media was replaced with either normal or Ca2+-depleted media and cultured for an additional 12 h. The resulting lysates were subjected to in vivo ubiquitination assay. (B) Phase-contrast micrographs of MCF10A and MDA-MB-231 cells grown under sparse or confluent conditions. Scale bar, 50 μm. (B, C) MCF10A and MDA-MB-231 cells were cultured under sparse and confluent conditions as in (B), and extracts were analyzed for the indicated proteins by Western blotting.
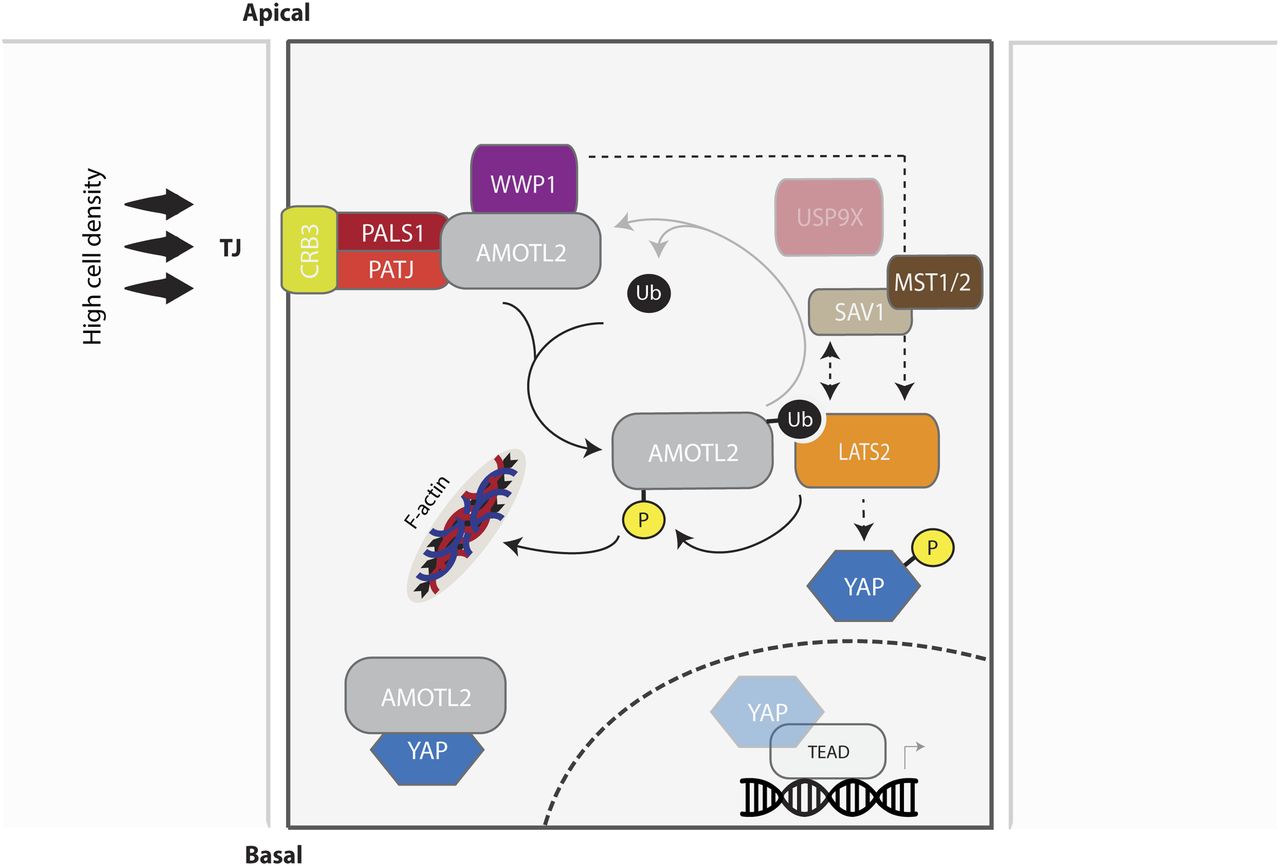
- Figure 8. Model for the regulation and role of WWP1 in the mono-ubiquitination of AMOTL2 and activation of LATS.
High cell density and the ensuing contact inhibition promotes assembly of the Crumbs complex, which recruits AMOTL2 to tight junctions. By virtue of its association with AMOTL2, WWP1 also preferentially localizes to the plasma membrane, where it becomes stabilized and actively mono-ubiquitinates AMOTL2. Ubiquitinated AMOTL2 then serves as a physical docking site for LATS2, which phosphorylates YAP to promote its cytoplasmic retention and degradation. WWP1 may also activate LATS2 independently from its ubiquitination effect on AMOTL2 (through a yet unidentified pathway), causing AMOTL2 phosphorylation and its displacement from filamentous actin (F-actin), thereby physically sequestering YAP within the cytosol. In summary, contact inhibition activates WWP1 and the ensuing bi-parallel mode of action of WWP1 hinders YAP nuclear entry and activity.









Supplementary Materials




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- No citing articles found.