



Abstract
The mechanisms underlying multipartite genome maintenance and its functional significance in extraordinary radioresistance of Deinococcus radiodurans are not well understood. The sequences upstream to parAB operons in chrII (cisII) and MP (cisMP) could stabilize an otherwise, non-replicative colE1 plasmid, in D. radiodurans. DnaA and cognate ParB proteins bound specifically with cisII and cisMP elements. The ΔcisII and ΔcisMP cells showed the reduced copy number of cognate replicons and radioresistance as compared with wild type. Fluorescent reporter–operator system inserted in chrI, chrII, and MP in wild type and cisII mutants showed the presence of all three replicons in wild-type cells. Although chrI was present in all the ΔcisII and ΔcisMP cells, nearly half of these cells had chrII and MP, respectively, and the other half had the reduced number of foci representing these replications. These results suggested that cisII and cisMP elements contain both origin of replication and parS-like functions and the secondary genome replicons (chrII and MP) are maintained independent of chrI and have roles in radioresistance of D. radiodurans.
Introduction
DNA replication and segregation are the highly coordinated macromolecular events required in the growth of any organism (Badrinarayanan et al, 2015). The origin of replication (oriC) where chromosomal replication in bacteria initiates is a well-conserved region on chromosomes and comprises of the non-palindromic repeats of 9-mer as DnaA boxes and 13-mer AT-rich repeats (Ogasawara et al., 1985; Marczynski & Shapiro, 1992; Messer, 2002). In Escherichia coli, DnaA, a replication initiation protein, first binds to the DnaA boxes which subsequently leads to the recruitment of replication initiation complex comprising of DnaB helicase, primase, and DNA polymerase III (Tougu & Marians, 1996; McHenry, 2011). Earlier, the tripartite genome segregation comprising (i) an origin-proximal cis-acting (parS-like) DNA sequences, (ii) parS binding proteins like ParB or its homologues, and (iii) the P-loop Walker ATPases such as ParA or its homologues has been reported to be the major mechanisms associated with the segregation of duplicated genome into daughter cells (Hayes & Barillà, 2006). Typically, ParB homologues first bind to the parS-like sequences and the interaction of ParA homologues to ParB-parS complex leads to either polymerization or depolymerization of ParA as required for DNA segregation (Hayes & Barillà, 2006; Gerdes et al, 2010; Reyes-Lamothe et al, 2012). Among different mechanisms of plasmid/chromosome segregation in bacteria, the diffusion-ratchet model is found to be the most acceptable (Vecchiarelli et al, 2010). Recently, it is shown that the duplicated chromosome segregates into daughter cells by the cumulative actions of different genome maintenance proteins viz. ParA-ParB, SMC (structural maintenance of chromosome)/MukB, topoisomerases, translocases, and nucleoid-associated proteins (Pióro & Jakimowicz, 2020). Furthermore, it has been observed that the initiation of DNA replication and segregation of duplicated chromosomes occur concurrently (Marczynski et al, 2019). In the majority cases where DNA replication and segregation have been studied, the parS-like sequences are found close to the origin of replication (ori) region. For instance, in Bacillus subtilis and Caulobacter crescentus, the primary parS sequences are located within 8–10-kb from oriC (Livny et al, 2007). Furthermore, the genome segregation proteins interact directly with DnaA in many bacteria (Murray & Errington, 2008; Marczynski et al, 2019). The organization and dynamics of nucleoid during cell growth have been studied in monopartite genome harboring rod- or crescent-shaped bacteria such as E. coli, B. subtilis, and C. crescentus (Esnault et al, 2007; Fisher et al, 2013; Le et al, 2013; Badrinarayanan et al, 2015; Dame & Tark-Dame, 2016) and cocci-like Streptococcus pneumonia (Kjos & Veening, 2017) and Staphylococcus aureus (Morikawa et al, 2006, 2019).
Recently, many bacteria have been reported with multipartite genome system comprising more than one chromosome and large plasmids (Misra et al, 2018). Studies on multipartite genome maintenance in bacteria are limited to Vibrio cholerae, Burkholderia cenocepacia, Rhodobacter sphaeiroides, and Shinorhizobium meliloti (Fogel & Waldor, 2005; Dubarry et al, 2006, 2019; Galardini et al, 2013). In these bacteria, the proteins of tripartite genome segregation encoded on the primary chromosome are phylogenetically similar to the chromosome of monopartite genome-harboring bacteria and found to be different from secondary genome components. Because the sizes of genomic replicons are different, how their replication and segregation are synchronized before cytokinesis would have occurred is intriguing. However, the real-time monitoring of DNA replication in V. cholerae a multipartite genome harboring (MGH) bacteria has shown that replication initiation of different replicons is staggered to terminate roughly at the same time (Rasmussen et al, 2007; Kemter et al, 2018).
Deinococcus radiodurans, a Gram-positive nonpathogenic bacterium characterized for extraordinary radioresistance, also harbors a multipartite genome comprising chromosome I (2,648,638 bp), chromosome II (412,348 bp), megaplasmid (177,466 bp), and small plasmid (45,704 bp) (White et al, 1999). Except small plasmid, all other replicons encode ParA- and ParB-like proteins in parAB operons. Earlier, Minsky and colleagues demonstrated that the multipartite genome of this bacterium is packaged in the form of a doughnut-shaped toroidal structure (Levin-Zaidman et al, 2003). Recently, the nucleoid dynamics has been monitored at different stages of growth in D. radiodurans and observed that the highly condensed nucleoid adapts multiple configurations during cell cycle progression, and the oriC in chrI remains radially distributed around the centrally positioned ter (terminus) sites (Floc’h et al., 2019). The characterization of ori and parS-like sequences in chromosome II (chrII) and megaplasmid (MP), and their localizations in nucleoid have not been reported. Here, we report that the direct repeats located upstream of parAB operons in chrII (cisII) and MP (cisMP) confers both ori and parS-like functions. Furthermore, we demonstrate that DnaA encoded on chrI interacts with these cis-elements of secondary genome components, whereas ParB of chrII (ParB2) and megaplasmid (ParB3) shows specific interaction with cognate cis-elements in vitro. Interestingly, both these cis-elements help in the maintenance of an otherwise suicidal plasmid, in D. radiodurans. The deletion of cisII (ΔcisII) and cisMP (ΔcisMP) leads to reduction in copy numbers of respective replicons and the partial loss of γ-radiation resistance as compared with wild type. These cis mutants produce an increased frequency of cells devoid of cognate replicons. The tetO-TetR-GFP–based fluorescent reporter–operator system (FROS) inserted near the putative origin of replication in chrI, chrII, and MP allowed us to localize these replicons in the nucleoid. Most ΔcisII and ΔcisMP cells showed the loss of chrII and MP, respectively, whereas there was no change in the wild-type pattern of chrI in these cells. These results suggested that cisII and cisMP confer both ori and parS-like functions, and DnaA encoded on the primary chromosome seems to regulate chrII and MP replication. Nearly no change in chrI localization pattern and its copy number in both the cis mutants, which has affected the maintenance of cognate replicons, further concluded that primary chromosome and secondary genome replicons are maintained independently in this bacterium.
Results
Sequences in chrII and MP predicted as putative “oriC” and parS-like sequences
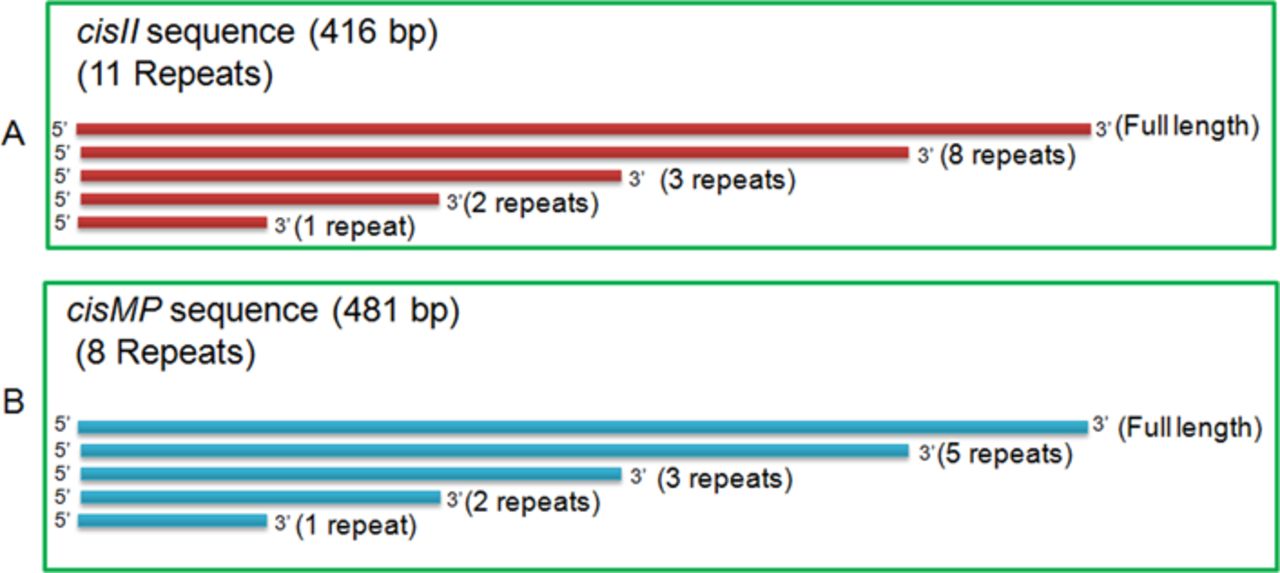
The upstream of parAB operons in chrII and MP contains repetitive sequences similar to the known bacterial “oriC” and parS-like cis-elements. The region spanning between 99 and 544 bp in chrII contain 11 iterons of 18 bp with consensus sequence T(G)CCACAAAGTGCCA(G)CAGG and GC content of 51.6% was named as cisII (Fig 1A). Similarly, the region spanning 177,446–417 bp ion MP contains eight iterons of 18 bp with consensus sequence C(A)CCGCAAAGGTG(A)TCGCTA and 3 iterons of 14 bp with consensus sequence TTTTGACCCCAAAT and GC content of 47% together named as cisMP (Fig 1B). In addition, both cisII and cisMP elements contain non-canonical putative DnaA boxes which were different from the 13 9-mer DnaA boxes (T(A/G)TA(T)TCCACA) present in the origin of replication of chrI in D. radiodurans and DnaA boxes in oriC of E. coli (Messer, 2002; Maurya & Misra, 2020 Preprint). Although the DnaA boxes started just upstream to 18-bp iterons, they continued into the iterons resulting to considerable overlap between both the elements. In addition, both the cis-elements contain AT-rich sequences, which are the known signatures of “ori-”like sequences needed for replication initiation in bacteria. In a previous study, three parS-like regions with consensus of GTTTC(A)G(A)C(A)GT(C)GG(T)A(G)AAC were mapped at 3.5° (segS1), 72.9° (segS2) and 231.8° (segS3) in chrI (Charaka & Misra, 2012). The predicted parS-like sequences in chrII and MP are found to be distinct from the conserved segS sequence of chrI. Thus, we found that cisII and cisMP elements have DnaA boxes similar to “oriC” feature and the iterons as parS-like sequences. The presence of the different “ori” and parS-like signatures in chrI from that of secondary genome replicons like chrII and MP indicated a strong possibility of independent maintenance of primary chromosome and secondary genome replicons in D. radiodurans.
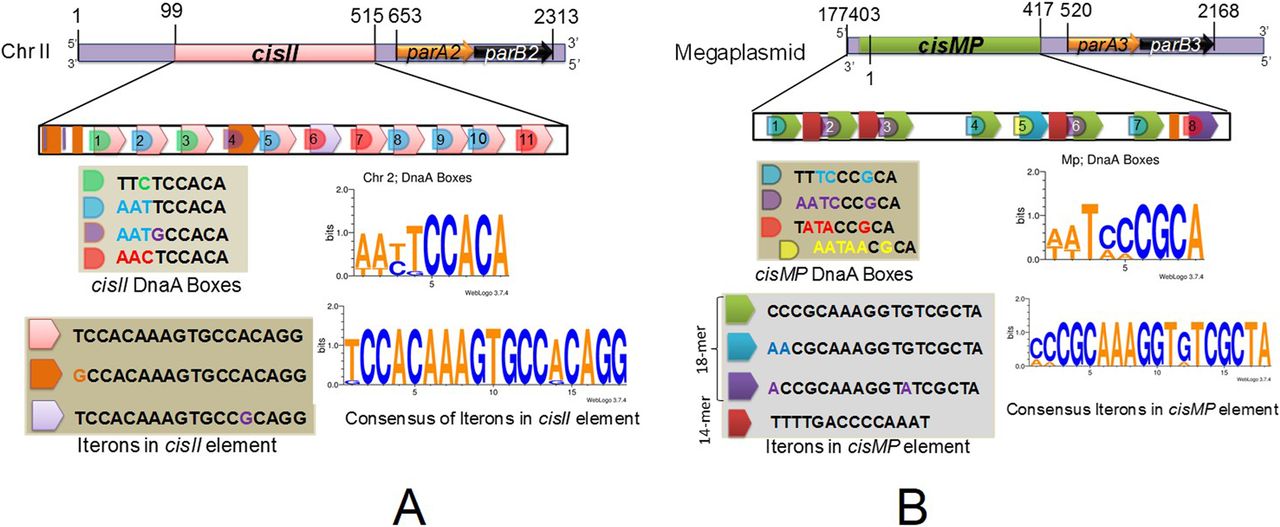
(A, B) The 99–554 bp region of chromosome II (A) and 177,446- to 417-bp region of megaplasmid (B) were analysed in silico for the structure of direct repeats and their sequence compositions of DnaA boxes and iterons.
cisII and cisMP elements help E. coli plasmid maintenance in D. radiodurans
The function of cisII and cisMP as origin of replication was investigated using an E. coli plasmid pNOKOUT (KanR) that was non-replicative in D. radiodurans. The pNOKCisII and pNOKCisMP mini replicons that contain cisII and cisMP elements, respectively, were created in pNOKOUT and their independent maintenance in D. radiodurans was monitored. The D. radiodurans cells harboring pNOKCisII or pNOKCisMP could multiply in the presence of kanamycin (6 μg/ml), which was not observed when these cells were transformed with pNOKOUT vector (Fig 2A and B). Furthermore, the independent maintenance of pNOKCisII or pNOKCisMP mini replicons was also ascertained in ΔrecA mutant of D. radiodurans and by recovering the intact plasmid DNA from the recombinant cells. We observed that the ΔrecA cells harboring pNOKCisII or pNOKCisMP replicons continued expressing kanamycin resistance like wild-type transformants of these plasmids (Fig 2C). The plasmids isolated from the wild-type transformants of D. radiodurans showed the typical patterns of original plasmid on an agarose gel and the release of the expected size inserts upon restriction digestion (Fig 2D). When we measured the copy number of these plasmids by qPCR, it was found to be 6.86 ± 0.71 for pNOKCisII and 12.15 ± 0.92 for pNOKCisMP. These results together suggested that pNOKCisII and pNOKCisMP mini replicons have been maintained independently in the D. radiodurans cell and not integrated into its genome, supporting the “ori” nature of cisII and cisMP elements in this bacterium. The cis-elements helping the non-replicative pNOKOUT plasmid to replicate in D. radiodurans, the possibility of these cis-elements working as parS-like sequences and thereby helping in the segregation of these mini replicons into daughter cells was hypothesized.
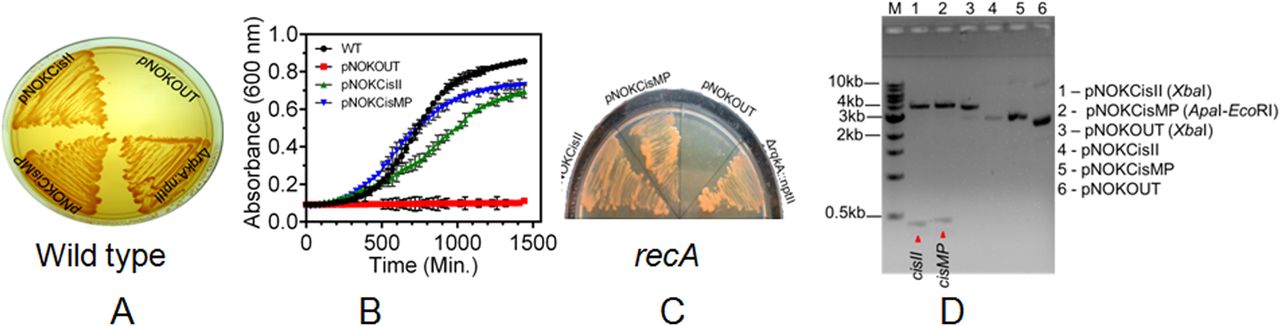
(A, C) The cisII and cisMP elements were cloned in E. coli cloning vector pNOKOUT, and the resultant plasmids pNOKcisII and pNOKcisMP were transformed into wild-type (A) and recA mutant (C). These transformants were grown in the presence of kanamycin (6 μg/ml) and growth was monitored on TGY agar plate with rqkA deletion mutant (a deletion mutant made by inserting kanamycin cassettes) as described in Rajpurohit & Misra (2010) as positive control and vector as a negative control. (B) The growth characteristic of these transformants grown in TGY broth supplemented with antibiotic was compared with wild-type cells grown in TGY broth without antibiotics (B). (D) The recombinant plasmids isolated from D. radiodurans cells growing in the presence of antibiotic were digested with restriction enzymes, and the release of cisII and cisMP fragments was analysed on an agarose gel and compared with empty vector (D).
cisII and cisMP elements provide sequence-specific interaction to DnaA
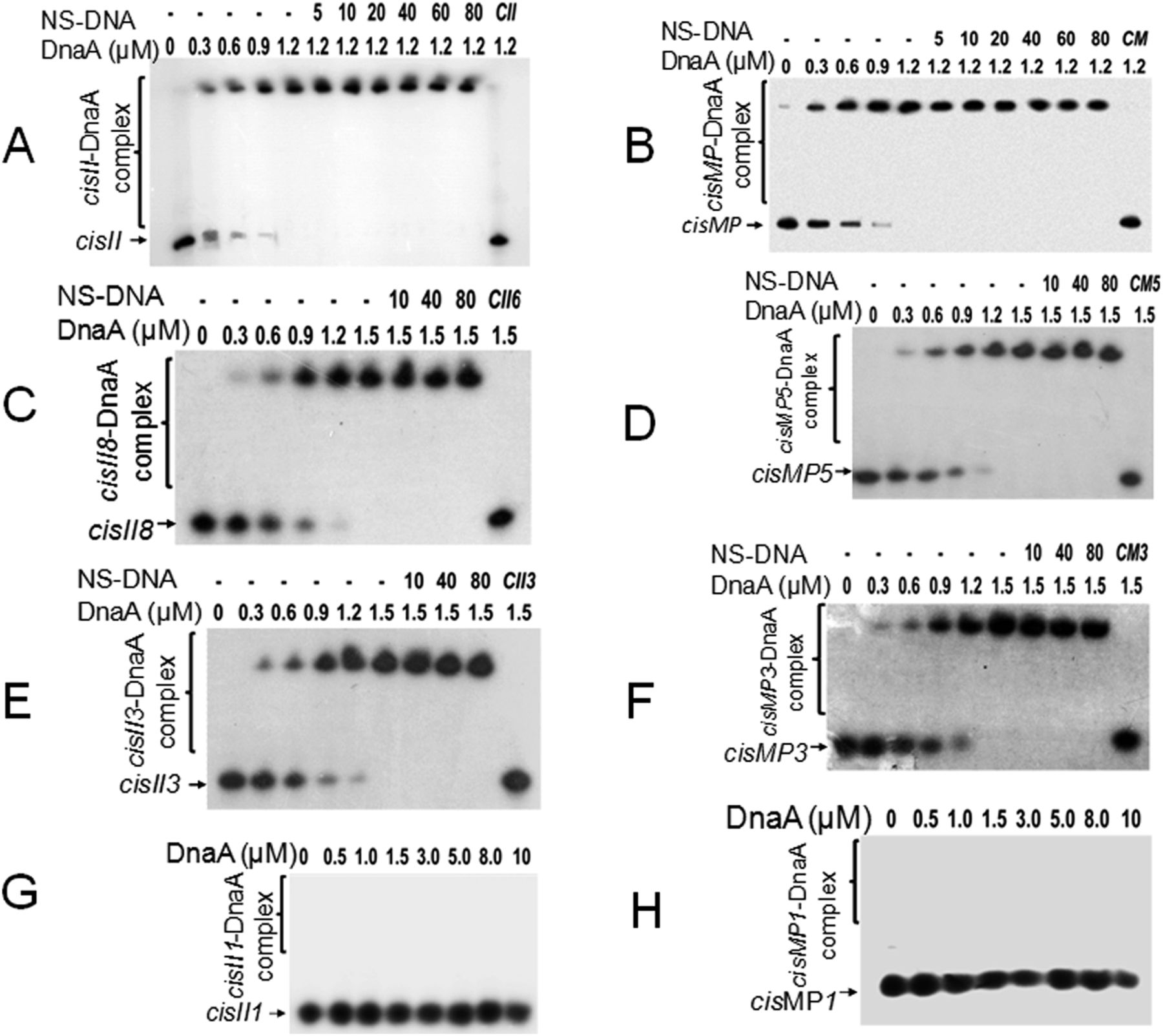
The presence of the known DNA replication initiation proteins such as DnaA; RctB of V. cholerae (Egan & Waldor, 2003); RepC homologues as known in Agrobacterium tumefaciens (Pinto et al, 2011), Brucella abortus (Pinto et al, 2012), Burkholderia pseudomallei (Holden et al, 2004), and S. meliloti (Cervantes-Rivera et al, 2011; Pinto et al, 2012); and RepN of B. subtilis (Tanaka et al, 2005) and S. aureus (Balson & Shaw, 1990) were searched on chrI, chrII, and MP of D. radiodurans. ChrII and MP do not encode for any of the known DNA replication initiators and DnaA homologue is encoded on chrI (data not given). Thus, the possibility of this DnaA interaction with cis-element of chrII and MP was checked. The recombinant purified DnaA (Fig S1A) showed sequence-specific interaction with both cisII (Fig 3A) and cisMP (Fig 3B) albeit with a different affinity. The affinity of DnaA varies as the number of repeats was reduced (Fig 3A–H and Table 1). As expected, the DnaA bound to either of the cis-elements remained unaffected even in the presence of an 80-fold higher molar concentration of nonspecific DNA (Fig 3). The formation of DnaA complex with cisII and cisMP containing several DnaA boxes suggests the “ori” nature of both the cis-elements in D. radiodurans.

(A, B, C) The recombinant DnaA (A), ParB2 (B), and ParB3 (C) were purified from E. coli using metal affinity chromatography.

(A, B, C, D, E, F, G, H) The radiolabelled full-length cisII containing 11 repeats (A) and cisMP containing eight repeats (B) as well as eight repeats (C), three repeats (E) and one repeat (G) of cisII (Fig S4A), and five repeats (D), three repeats (F), and single repeat (H) of cisMP (Fig S4B) were incubated with the increasing concentration of DnaA. A saturating concentration of the DnaA-DNA ratio was chased with increasing molar ratio of non-specific DNA (NS-DNA) and products were analysed on native PAGE. The fractions of DNA bound with proteins were quantified densitometrically and plotted as a function of protein concentration using GraphPad Prism 6. The Kd for the curve fitting of individual plots was determined and given in Table 1.
The dissociation constant (Kd) value of DnaA interaction with different repeats of cisII and cisMP elements.
ParB2 and ParB3 show specific interaction with cognate cis-elements
Because both the cis-elements have iterons as a potential parS-like sequences and ParB homologues are known to bind parS-like sequences, the interactions of ParB2 and ParB3 with their cognate cis-elements were monitored. The radiolabelled cisII and their repeat variants (Fig 4A–D) as well as cisMP and their repeat variants (Fig 4E–H) were incubated with purified recombinant ParB2 (Fig S1B) and ParB3 proteins (Fig S1C), respectively, and the products were analysed on native PAGE. The ParB2 and ParB3 showed higher affinity with full-length cisII (Kd = 0.40 ± 0.007 μM) and cisMP (0.59 ± 0.05 μM), respectively (Fig 4A and E). The affinity of ParBs to cognate cis-element was nearly same up to three repeats and reduced when the number of repeats becomes less than three (Fig 4C and G and Table 2). These proteins are sparingly bound to DNA substrates with less than two repeats (Fig 4D and H). The binding of ParBs to the cognate cis-elements was sequence specific as it was not affected in the presence of up to 100-fold higher molar concentration of nonspecific DNA (Fig 4A–C and E–G). Furthermore, it was noticed that the full-length cisII and cisMP bound ParBs migrated under electrophoresis with peculiar patterns that are normally seen when the protein binding causes a structural change like bending in the DNA. These findings suggested that both cisII and cisMP elements have a specific region for interaction with respective ParB2 and ParB3, which might function as parS-like sequence in the faithful segregation of cognate replicons.
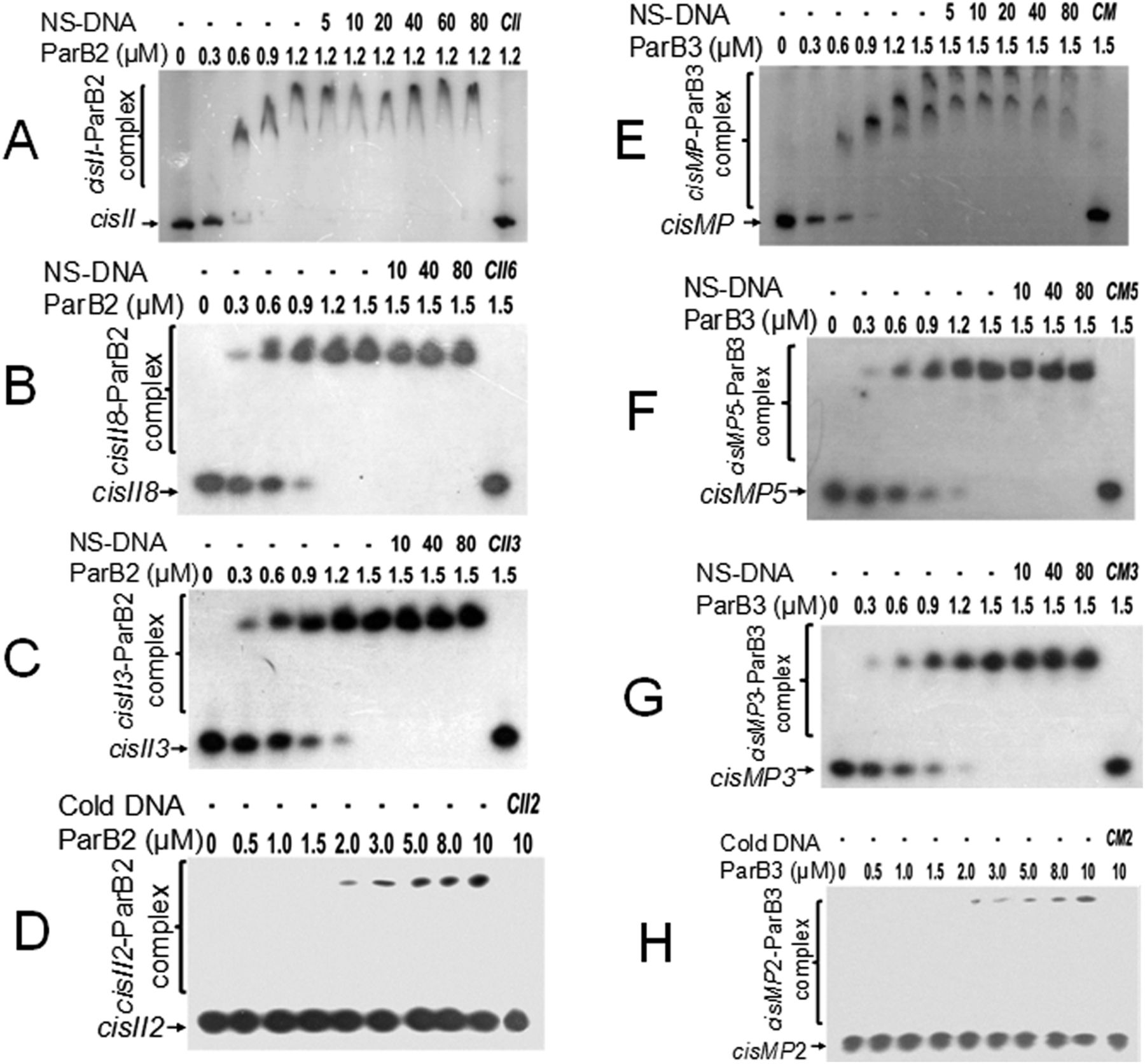
(A, B, C, D) The radiolabelled full-length cisII containing 11 repeats (A), 8 repeats (B), 3 repeats (C), and 2 repeats (D) (Table S2) were incubated with the increasing concentration of ParB2. (E, F, G, H) Similarly, the radiolabelled full-length cisMP containing eight repeats (E), five repeats (F), three repeats (G), and two repeats (H) were incubated with the increasing concentration of ParB3. A saturating concentration of DNA–protein ratio was chased with increasing molar ratio of non-specific DNA (NS-DNA) and products were analysed on native PAGE. The fractions of DNA bound with proteins were quantified densitometrically and plotted as a function of protein concentration using GraphPad Prism 6. The Kd for the curve fitting of individual plots was determined and given in Table 2.
The dissociation constant (Kd) value of ParB2 and ParB3 interaction with repeats variants of cisII and cisMP elements, respectively.
cisII and cisMP deletion affected cognate replicons copy number and their segregation
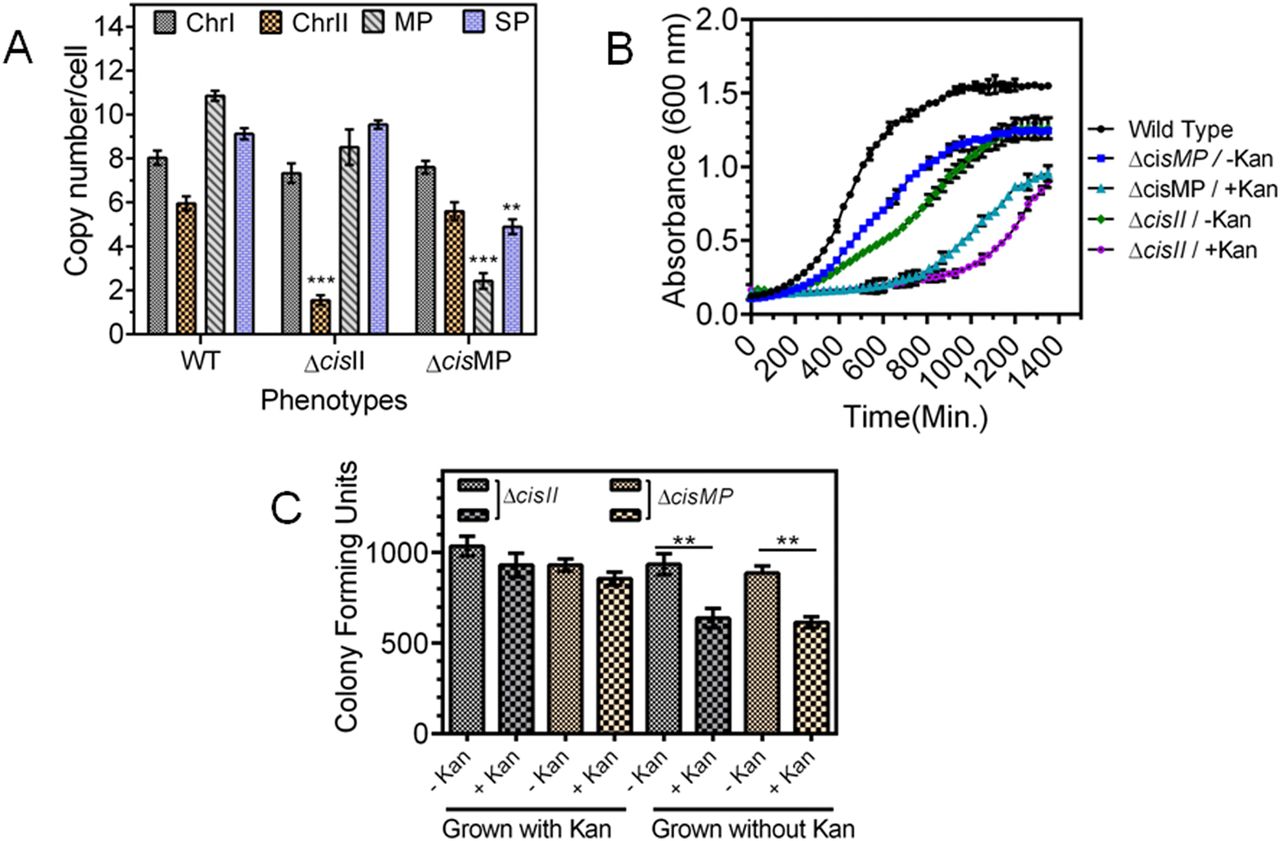
To understand the essentiality of chrI, chrII, and MP, the possibility of deleting cisII, cisMP, and putative ori of chrI (oriCI) was explored. We could not obtain the deletion mutant of oriCI. However, the homozygous replacement of cisII (Fig S2A and B) and cisMP (Fig S2A and C) with an expressing cassette of nptII was achieved, and these cells were viable in both liquid medium and agar plates. Therefore, the effects of cisII and cisMP deletion on the stability of cognate replicons and their genome copy numbers were determined in mutant cells and compared with wild type. A significant reduction in the copy number of chrII and MP was observed in ΔcisII and ΔcisMP cells, respectively, but the copy number of chrI did not change in any of the mutant backgrounds (Fig 5A). For instance, the average copy number of chrII reduced from 5.95 ± 0.32 to 1.55 ± 0.22 and MP was reduced from 10.85 ± 0.24 to 8.52 ± 0.81 in ΔcisII mutant. Similarly, the average copy number of MP was reduced from 10.85 ± 0.24 to 2.42 ± 0.35 in ΔcisMP mutant (Fig 5A). The deletion of cisMP did not affect the copy number of either chrI or chrII when compared with wild type. The reduction in ploidy of secondary genome replicons in deletion mutant of cognate cis-elements could be accounted for its effect on the replication of these replicons. However, a possibility of these deletions affecting DNA segregation in dividing cells cannot be ruled out. This possibility was checked by monitoring the frequency of cells conferring KanR (expressed on ChrII or MP) when grown in the absence and presence of kanamycin. In general, the growth of cis mutants was affected as compared with wild type and reduced further when grew in the presence of kanamycin as compared with its absence (Fig 5B). Furthermore, the ΔcisII and ΔcisMP mutants were grown for 14 h (log phase) in the presence and absence of kanamycin and total CFUs were measured in TGY agar plate in the presence and absence of antibiotic. Results showed that KanR CFU was reduced nearly half (−Kan vs +Kan) in both the mutants when they were grown without selection pressure, whereas there was no change in KanR CFU (−Kan vs +Kan) when these mutants were maintained with antibiotic selection (Fig 5C). Because the antibiotic is expressed on respective replicons, these results indicated that all daughter cells grown without selection pressure might have not inherited cognate replicon in cis mutant. Therefore, a possibility of chrI alone supporting the normal growth of this bacterium was suggested. Similar studies have been reported in B. subtilis and Mycobacterium tuberculosis (Hassan et al, 1997; Qin et al, 1999; Richardson et al, 2016, 2019). These results together suggested that cisII and cisMP elements seem to carry functions for controlling both DNA replication and segregation in D. radiodurans. Furthermore, the status of chrII and MP in cis mutants was examined microscopically.

(A) Schematic representation of strategy for replacement of cis-elements with the expressing cassette of kanamycin and position of primers used for diagnostic PCR (A). (B, C) The recombinant cells expected to have cisII (B) and cisMP (C) deletions grown several generations under selection pressure. (A) The genomic DNA was prepared, and PCR amplification was carried out by diagnostic PCR using primer as shown in (A). Products were analysed on 1% agarose gel.

(A) The exponentially growing D. radiodurans R1 (WT) and its cisII (ΔcisII) and cisMP (ΔcisMP) deletion mutants were used for the determination of copy number of each replicon (A). (B) Both wild type and cis mutants of D. radiodurans were checked for growth in the presence and absence of kanamycin under normal conditions (B). (C) These mutants were maintained in the presence (+Kan) and absence (−Kan) of antibiotics as indicated on x-axis, and the CFUs in the absence and presence of kanamycin were monitored and plotted on y-axis (C).
cisII and cisMP deletions affected the maintenance of cognate genomic replicons
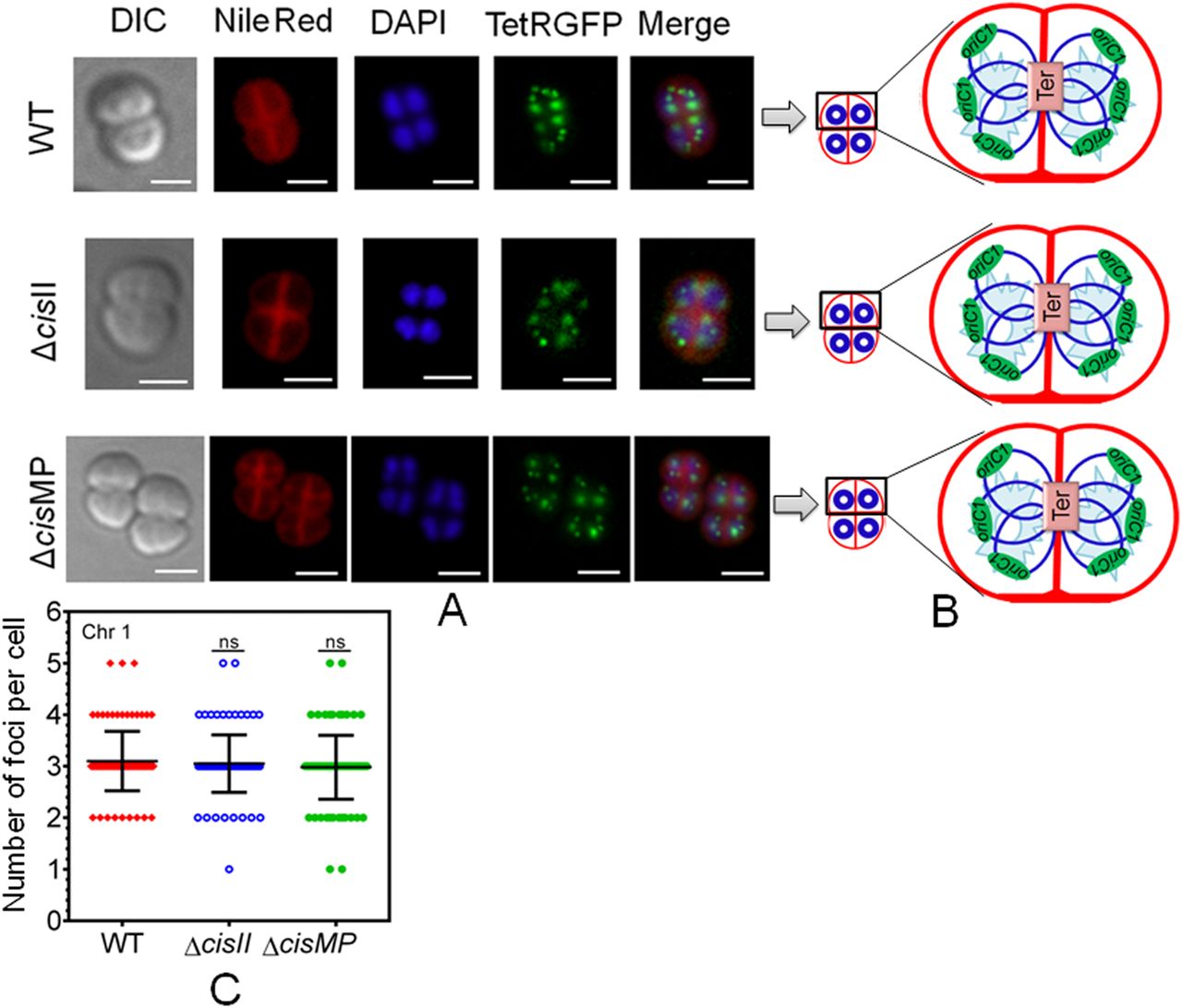
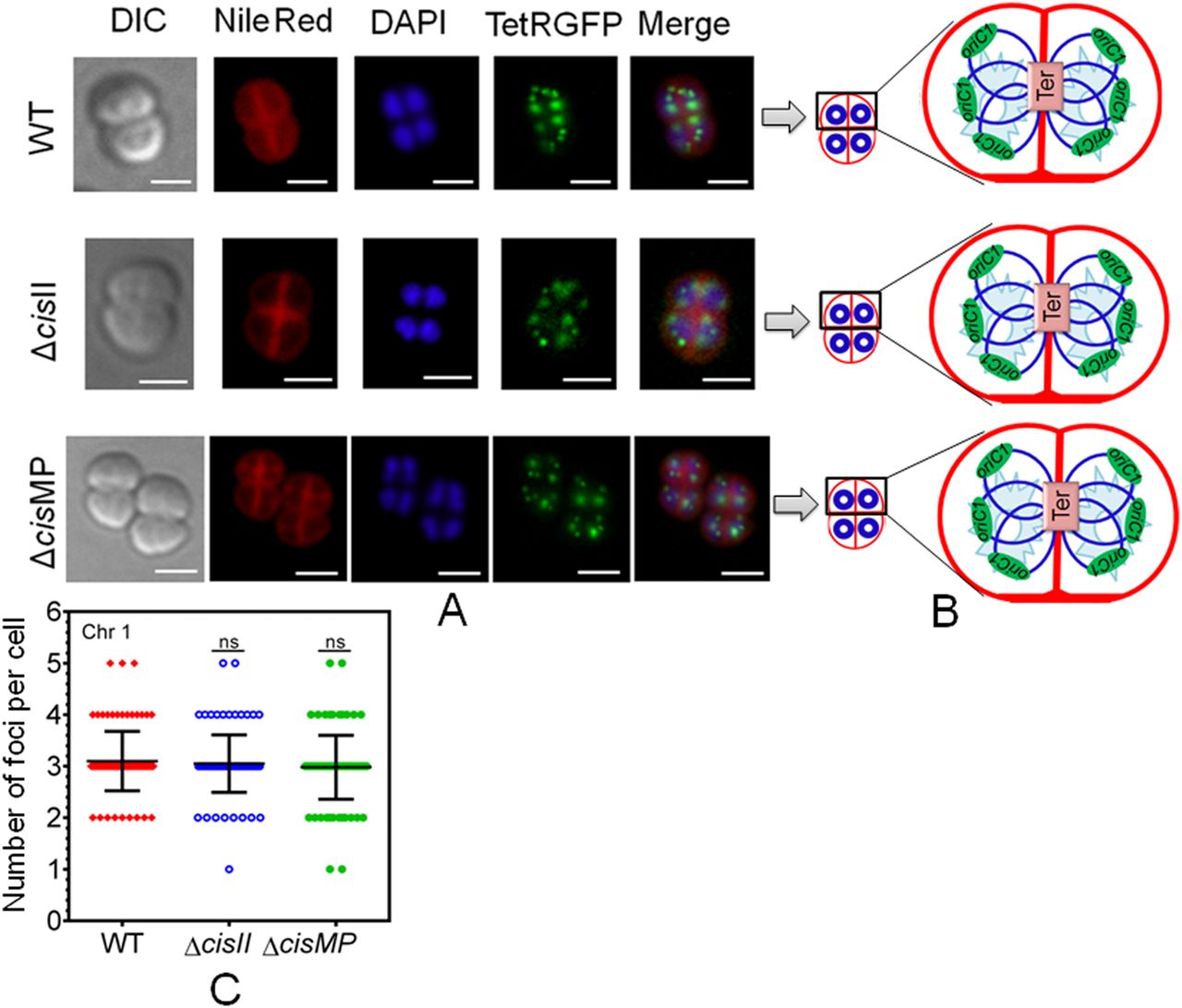
The multipartite genome system, ploidy, and packaging of complete genetic materials into a doughnut-shaped toroidal nucleoid are considered to be the remarkable cytogenetic features in D. radiodurans (Harsojo & Matsuyama, 1981; Levin-Zaidman et al, 2003; Minsky et al, 2006). However, the organization of genomic replicons in toroidal nucleoid has not been demonstrated. In this work, we developed an FROS approach for the chrI (Fig S3A), chrII (Fig S3B), and MP (Fig S3D) in the wild-type, and ∆cisII- and ∆cisMP-mutant cells as described in the Materials and Methods section (Fig S3C and E). The exponentially growing cells were stained with Nile red and DAPI and imaged under a fluorescence microscope. Interestingly, the localization pattern of chrI in the ΔcisII and ΔcisMP mutants and wild-type cells were similar (Fig 6A). The average number of foci for chrI per cell was ∼3, which were localized away from the septum and seems to be radially distributed throughout the cytoplasm in both wild type and ΔcisII and ΔcisMP cells (Fig 6B and C). A nearly similar pattern of chrI localization has been recently demonstrated in D. radiodurans, where the number of foci per nucleoid was shown to be in the range of 2.25–4.44 (Floc’h et al, 2019). However, when the localization pattern of chrII (Fig 7A) and MP (Fig 8A) was observed under microscope, the average number of foci for chrII (Fig 7A–C) and MP (Fig 8A–C) was found to be 2.09 and 1.1 per cell, respectively. These were too located away from the septum and spanned throughout the nucleoid in wild type. However, in the case of cis mutant, the localization pattern of chrII and MP was different. For instance, chrII localization in nucleoid was altered in ΔcisII but not in ΔcisMP and vice versa. Both these cis mutants produced a population of cells that did not contain the cognate replicon. Because the qPCR study has shown the highest number (10.85) of copies of megaplasmid per cell, the single GFP foci observed through FROS is intriguing but informative. Similar results have been reported in other limited copy plasmids such as RK2, R1, etc. in E. coli (Weitao et al, 2000; Pogliano et al, 2001). The decreasing number of GFP foci from 3 to 4 in ChrI to one in MP agreed with their size difference and if that has affected resolution cannot be ruled out.

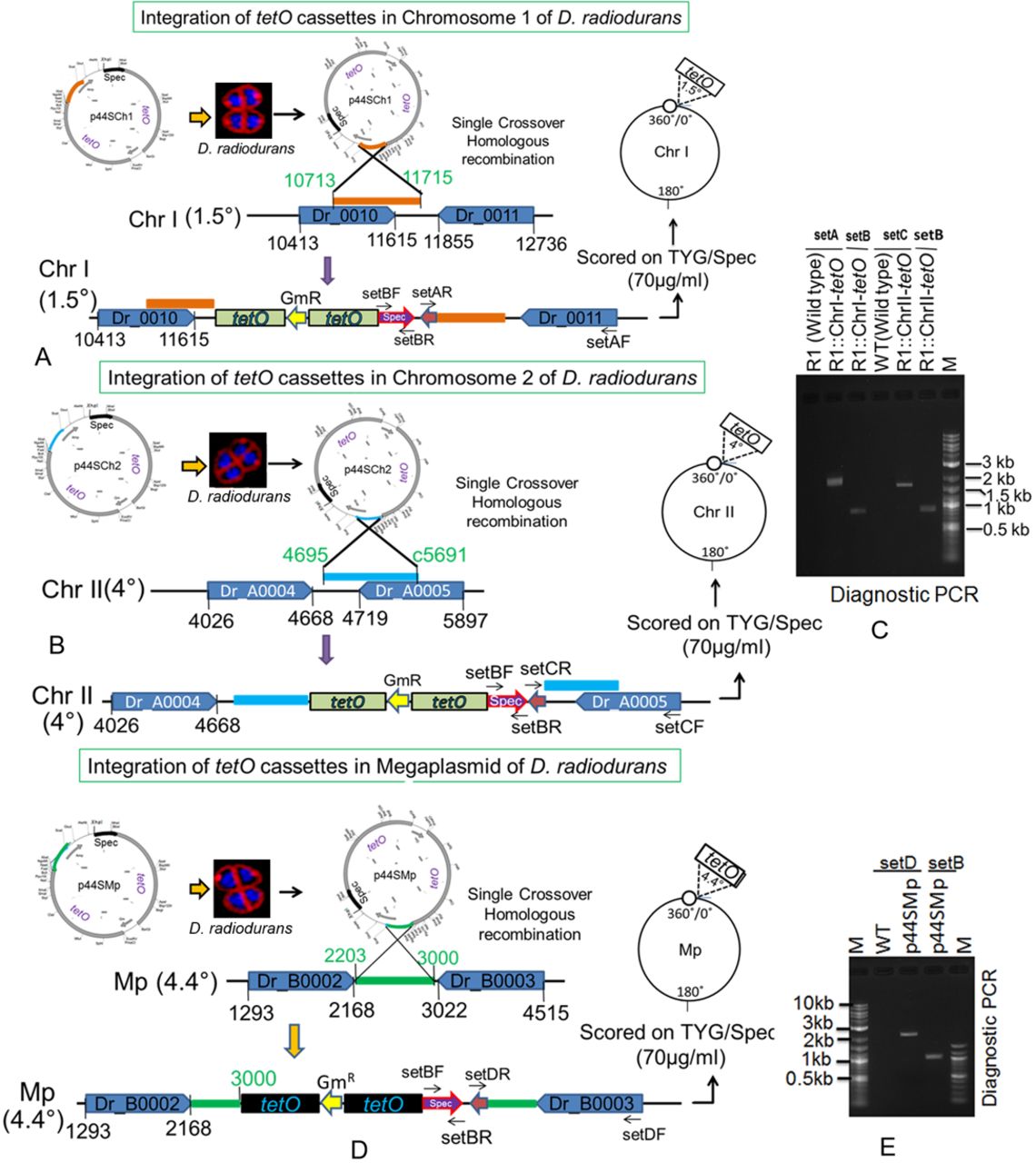
(A, B, D) The tetO cassette from was inserted at 1.5° in chromosome I (A), at 4° in chromosome II (B) and at 4.4° in megaplasmid (D) through homologous recombination. (C, E) Insertion of tetO cassettes into respective positions was confirmed by diagnostic PCR using plasmid sequence-specific primers (C, E).
Chromosome I was tagged with tetO/TetR-GFP–based fluorescent reporter–operator system in both wild type (WT) and cis mutants (ΔcisII and ΔcisMP) as described in the Materials and Methods section and grown exponentially. (A) These cells were stained with Nile Red and DAPI and visualized microscopically in differential interference contrast, TRITC (Nile Red), DAPI (DAPI), and FITC (GFP) channels and images were merged (Merge) (A). The scale bar of 1 µm is used in each figure. (B) The schematic diagrams showing the foci position with respect to nucleoid and septum are presented for better clarity (B). (C) Data shown are from a single tetrad where most of these cells show a similar pattern as quantified from ∼200 cells (C).

Chromosome II was tagged with a tetO-TetR-GFP–based fluorescent reporter–operator system in both wild type (WT) and cis mutants (ΔcisII and ΔcisMP) as described in the Materials and Methods section and grown exponentially. (A) These cells were stained with Nile Red and DAPI and visualized microscopically in differential interference contrast, TRITC (Nile Red), DAPI (DAPI), and FITC (GFP) channels and images were merged (Merge) (A). The scale bar of 1 µm is used in each figure. (B) The schematic diagrams showing the foci position with respect to nucleoid and septum are presented for better clarity (B). (C) Data shown are from a single tetrad where most of these cells show a similar pattern as quantified from ∼200 cells (C).
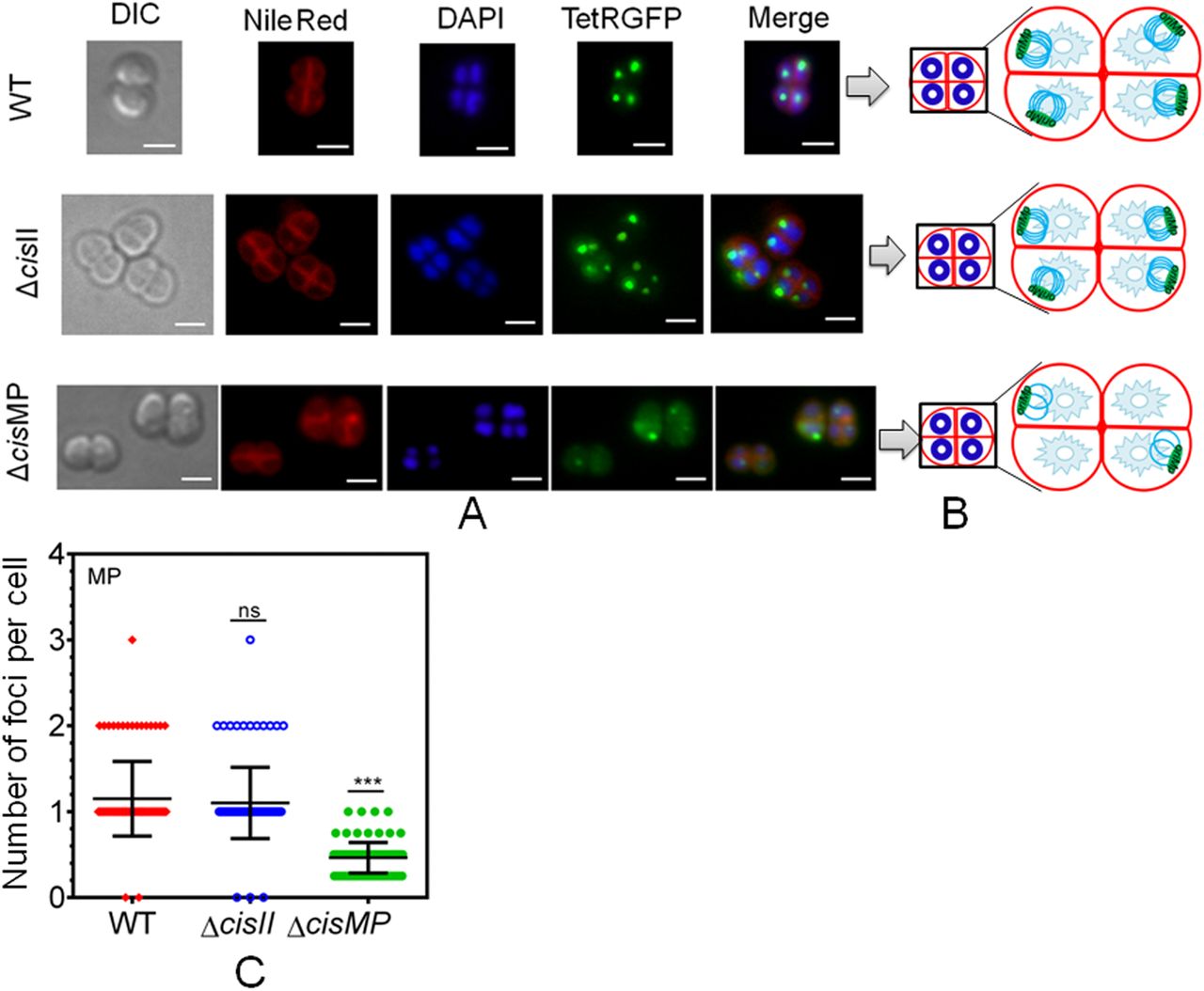
The megaplasmid was tagged with a tetO-TetR-GFP–based fluorescent reporter–operator system in both wild type (WT) and cis mutants (ΔcisII and ΔcisMP) as described in the Materials and Methods section and grown exponentially. (A) These cells were stained with Nile Red and DAPI and visualized microscopically in differential interference contrast, TRITC (Nile Red), DAPI (DAPI), and FITC (GFP) channels and images were merged (Merge) (A). The scale bar of 1 µm is used in each figure. (B) The schematic diagrams showing the foci position with respect to nucleoid and septum are presented for better clarity (B). (C) Data shown are from a single tetrad where most of these cells show a similar pattern as quantified from ∼200 cells (C).
Furthermore, we studied the population dynamics of cells for the presence and absence of chrI, chrII and MP in wild type and cis mutants. The majority of wild type cells showed ≥3 foci corresponding to chrI, 2 foci for chrII and single focus for MP (Fig 9A). However, the number of cells with ≥3 foci corresponding to chrI in both the cis mutants was nearly similar to wild type. For chrII, these were nearly the same in cisMP mutant and wild type with two foci in each. However, in cisII mutant, there was hardly any cell containing two foci and all the population was divided almost equally into cells with a single focus or no focus (Fig 9B). Similarly, wild-type and cisII mutant showed a nearly similar number of foci corresponding to MP, which was different in cisMP mutant. For instance, the cisMP mutant showed nearly half of the cells with a single focus and the other half population did not show foci corresponding to MP (Fig 9C). These results highlighted the roles of cisII and cisMP as “ori” and parS-like functions in the secondary genome replicons of D. radiodurans. Furthermore, the FROS-based localization provided the first evidence that both primary and secondary genome replicons are the part of the compact nucleoid and these replicons seem to be independently maintained in this multipartite genome-harboring bacterium.
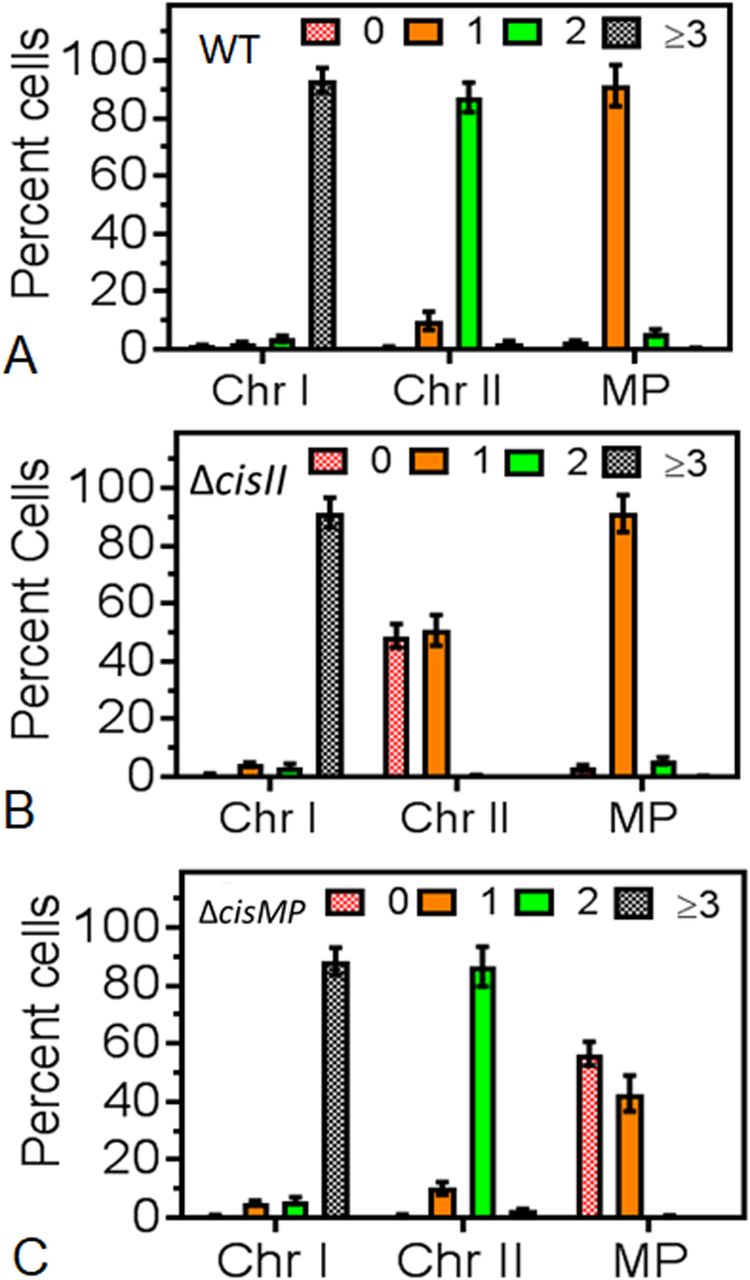
(A, B, C) The chromosome I (ChrI), chromosome II (ChrII and megaplasmid [MP]) of wild type (A), cisII (B), and cisMP (C) mutant of D. radiodurans were tagged with GFP-based fluorescent reporter–operator system and cells were imaged under fluorescence microscopy as described in Figs 6–8. Nearly 200 cells of each type were analysed for the absence (0) and the presence of varying numbers (1, 2, ≥ 3) of foci corresponding to all three replicons. Data presented here are mean ± SD (n = 200).
cisII and cisMP mutants showed reduction in γ-radiation resistance
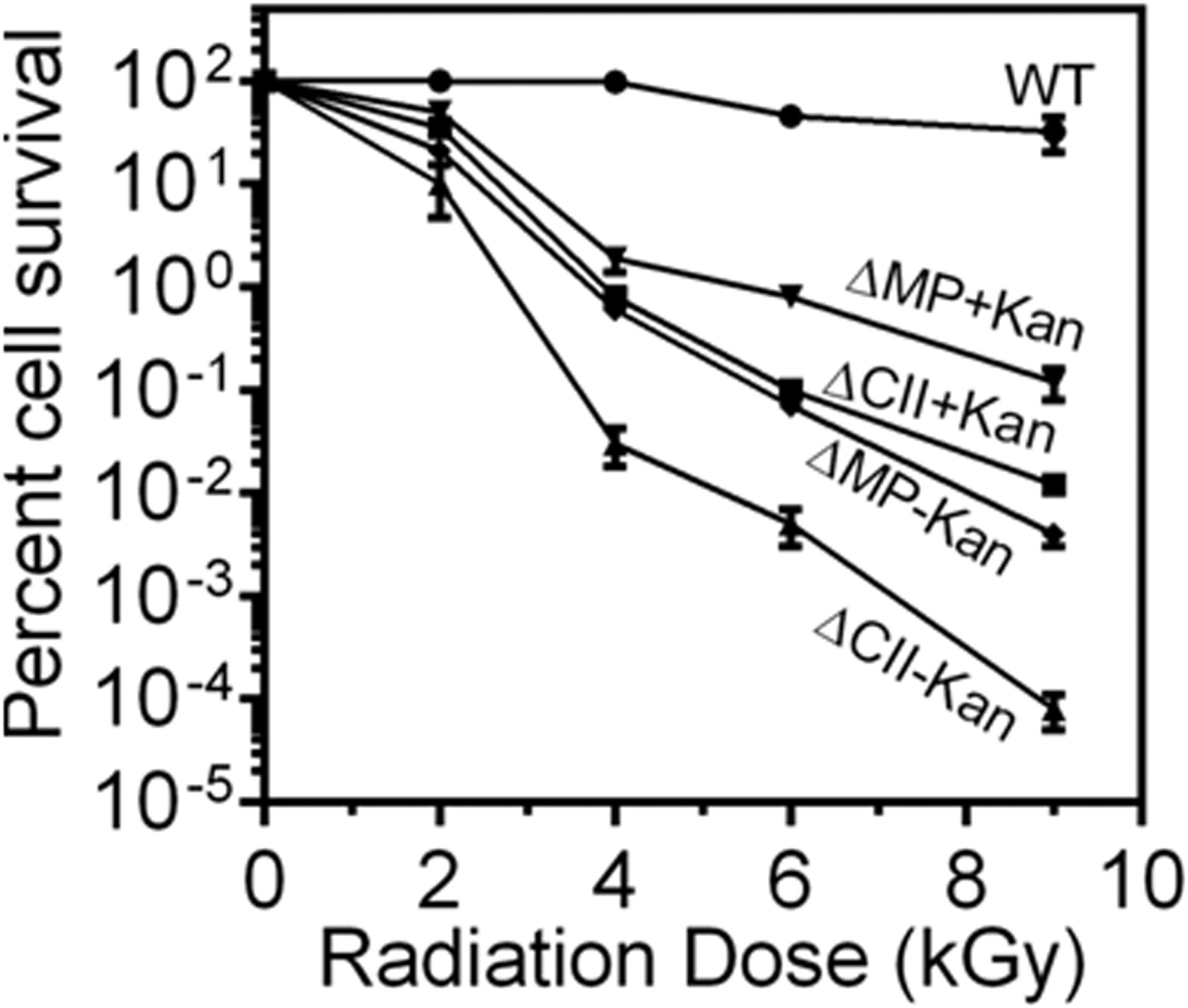
The effect of cisII and cisMP deletion on γ radiation response of D. radiodurans was examined. The ∆cisII mutant was more sensitive to γ radiation than ∆cisMP mutant (Fig 10). Furthermore, the mutants that were maintained without antibiotic selection showed nearly twofold higher sensitivity to γ radiation as compared with those grown with antibiotic (Fig 10). The loss of γ radiation resistance in cis mutants that apparently had shown the reduction in copy number in nearly half of the population, and the other half were anucleated for cognate replicons, suggested the role of chrII and MP in radioresistance of this bacterium. Earlier, it was shown that the reduction in copy number of chrII and MP in ΔparA2ΔparA3 double mutants has affected γ radiation response in D. radiodurans (Maurya et al, 2019a). The mechanisms responsible for the underlying loss of radioresistance in cells that have either a reduced copy number or the complete loss of chrII and MP are not understood. However, these replicons encode a large number of proteins including PprA (Narumi et al, 2004), DR_A0282 (Das & Misra, 2011), DR_A0074, and DR_A0065 (Ghosh & Grove, 2006) on chrII, and DR_B0100 (Kota et al, 2010), DR_B0090, and DR_B0091 (Desai et al, 2011), which have been characterized for their roles in DNA repair and radioresistance (Misra et al, 2013). Therefore, the reduction in the copy number or the absence of these proteins in cis mutants if have affected the radioresistance of D. radiodurans cannot be ruled out. These results suggested that chrII and MP replicons, which are perturbed in cisII and cisMP mutants, contributes to the radioresistance in this bacterium.

The γ radiation response of wild type (WT) and its cisII (ΔCII) and cisMP (ΔMP) mutants grown in the presence (+Kan) and absence (−Kan) of kanamycin was monitored. Data presented here are mean ± SD (n = 6) from the reproducible experiments.
Discussion
D. radiodurans, an extraordinary radioresistant bacterium is characterized for its peculiar cytogenetic features like the multipartite genome, ploidy, and tetrad cell morphology. Mechanisms underlying DNA replication and genome segregation have not been studied in detail in this bacterium, and very limited information is available in other MGH bacteria such as V. cholerae, R. sphaeroides, S. meliloti, and D. radiodurans (Fogel & Waldor, 2005; Charaka & Misra, 2012; Dubarry et al, 2019; Maurya et al, 2019a). Recently, the nucleoid dynamics and radial positioning of ori and ter in chrI of D. radiodurans have been demonstrated as a function of cell cycle (Floc’h et al, 2019). However, the organization of the secondary genome replicons in the nucleoid and their stable inheritance in dividing cells under normal and γ radiation stressed conditions had not been studied. This study has brought forth some evidences to suggest that the direct repeats present upstream to parAB operon in chrII (cisII) and MP (cisMP) function like “ori” and parS-like elements in D. radiodurans. By using FROS approach, we demonstrated the radial localization of chrI, chrII, and MP in the nucleoid and away from septum. The search of the homologues of RepC encoded on secondary genome element in A. Tumefaciens, B. abortus, B. pseudomallei, and S. Meliloti and RctB in V. cholerae did not pick up these proteins in the chrII and MP in D. radiodurans (data not shown). However, chrI encodes the DnaA homologue of E. coli. This does not rule out the possibility of some uncharacterized replication initiators for secondary genome elements in this bacterium.
We used DnaA and ParBs as probes for ori and parS-like sequences and confirmed that cisII and cisMP elements have both DnaA boxes and iterons. Our results demonstrating the (i) conversion of a non-replicative plasmid into a replicative one in the presence of these cis-elements, (ii) reduction in copy number of cognate replicon in absence of these elements, and (iii) an increased frequency of cells devoid of chrII and MP in respective deletion mutant, together supported the hypothesis that both cisII and cisMP elements play the important roles in the regulation of DNA replication and segregation functions in this bacterium. The presence of both “ori” and parS-like sequences in close vicinity, if helps the cells to differentially regulate the growth phase dependent interaction of replication complex with ori region and segregation complex with parS-like region cannot be ruled out. Earlier, the presence of parS-like sequences in proximity to “oriC” in their chromosomes and the molecular crosstalk between the components of the macromolecular events like replication and segregation has also been reported in some bacteria. For instance, in B. subtilis, Soj (a ParA homologue) directly interacts with DnaA proteins and regulates the replication at oriC (Scholefield et al, 2011). In C. crescentus, DnaA controls chromosome segregation in ParA dependent manner as well as by binding directly to the parS site (Marczynski et al, 2019). Although both the chromosomes in V. cholerae encode separate replication and segregation components (Egan & Waldor, 2003), the interdependent regulation of these two processes has been observed. For instance, ParA1, a genome segregation protein, encoded on chrI in V. cholerae stimulates DNA replication through its direct interaction with DnaA while ParB2, a parS binding protein, encoded on chrII showed inhibition of replication initiation in chrI (Kadoya et al, 2011). On contrary, ParB2 stimulates the replication of chrII by decreasing RctB binding to inhibitory DNA sequences adjacent to the oriII in V. cholerae (Pal et al, 2005; Venkova-Canova et al, 2013). In D. radiodurans, the interaction of segregation proteins with replication initiation proteins (DnaA and DnaB) and an arrest of genome segregation affecting DNA replication have been demonstrated (Maurya et al, 2019b; Maurya & Misra, 2020 Preprint). Although secondary genome elements in D. radiodurans do not encode for any of the known DNA replication initiation proteins, and cisII and cisMP are being implied to regulate DNA replication through DnaA, the possibility of some additional uncharacterized mechanism(s) for the regulation of replication in chrII and MP cannot be ruled out.
Furthermore, we tried to distinguish the replication and segregation functions of these cis-elements in vivo. We could delete cisII and cisMP from cognate genome elements but not oriCI from chrI of this bacterium and these cells were viable under normal conditions indicating a strong possibility that chrII and MP replicons are not required for the normal growth in this bacterium. Microscopically, we observed that only nearly half of the populations of ΔcisII and ΔcisMP mutants lack cognate replicon indicating a possibility of functional redundancy of these cis-elements in regulation of DNA replication in chrII and MP. Because chrII was designated as chromosome because of the presence of certain essential genes as chromosomal markers, nearly no effect of cisII deletion on survival of D. radiodurans is intriguing. However, the growth rates of these mutants under normal conditions and in the absence of antibiotic selection were different from the wild type. Results supported the hypothesis where we observed (i) the reduction in copy number of cognate replicons in mutant grown under antibiotic selection and (ii) the sizable cell populations of mutants did not have cognate replicons as monitored by the absence of antibiotic resistance encoded on them as well as microscopically. We argued that cis mutants, which were maintained without antibiotic selection may have grown as the heterogeneous population having cells with and without cognate replicons. When these are selected for KanR, nearly half of the population was found to be KanS (Fig 5B and C). Presumably, these were the ones that have survived with the help of chromosome I and were devoid of secondary replicons because of segregation defects. Although mutational analysis of these repeats and their interaction with cognate ParBs has provided a strong evidence on the role of these elements in genome segregation, loss of chrII and MP in nearly 50% population of cisII and cisMP mutants, respectively, also supports the role of these cis-elements in genome segregation.
Furthermore, the FROS results where 2–3 cells in tetrad were devoid of foci in respective replicons supported the argument of segregation defect in cells lacking cis-elements. Survival of other three cells in tetrad without replicons conferring antibiotic resistance under selection pressure is intriguing and a strong possibility of these supported by the neighbouring antibiotic-resistant cells cannot be ruled out. The careful monitoring of the localization pattern of chrI, chrII, and MP in nucleoid of wild type and cis mutants yielded very interesting results. The localization pattern of chrI was the same in both wild type and cisII and cisMP mutants, and agrees with the recent findings reported independently (Floc’h et al., 2019). Also, there was no change in the ploidy status of chrI in mutant cells. On the other hand, the localization pattern of chrII and MP in the nucleoid as well as the ploidy status of these replicons in cells deleted for cisII and cisMP elements, respectively, were grossly altered. Thus, we provided evidence that all three replicons are packaged in a single nucleoid and the “ori” region of these replicons is excluded from the membrane-proximal region of the cell. The anchoring of the chromosome to membrane-proximal region through SMC proteins has been reported in other bacteria (Ben-Yehuda et al, 2005; Thanbichler & Shapiro, 2006; Yamaichi et al, 2012). It would be worth mentioning that ParAs encoded on ChrII and MP are found to be functionally redundant (Maurya et al, 2019a), while ParB2 and ParB3 are not (Maurya et al, 2019b), and now cis-elements are differentially regulating both replication and segregation of cognate replicons in this bacterium.
In conclusion, for the first time, we have characterized the ori and parS-like regions in cisII and cisMP elements in chrII and MP and showed that these cis-elements regulate replication and segregation functions in an MGH bacterium D. radiodurans. Furthermore, we provided evidence that the maintenance of these secondary replicons is independent of the primary chromosome, yet the chrI, chrII, and MP are packaged in form of a compact nucleoid. Last, this study also provides evidence that the secondary genome replicons have roles in the extraordinary radioresistance of this bacterium. Recently, many bacteria have been discovered with a multipartite genome system, and surprisingly the most of these are pathogenic/inhabitant to some forms of life. Regulation of origin of replication, incompatibility and copy number as understood by taking E. coli as model system, would be worth revisiting in the context of ploidy and multipartite genome system as known in many MGH bacteria. This study may serve as an example and provide starting materials for answering many of these questions under newly born multipartite genome biology in bacteria. Apart from this, the independent studies would be undertaken to understand (i) the dynamics of the multipartite genome during γ radiation recovery and normal cell cycle regulation in this bacterium, and (ii) DNA damage responsive territorial change of genomic replicons in cells recovering from γ radiation exposure and its impact on genome functions in D. radiodurans. The presence of chrII and MP in certain population of cisII and cisMP mutants, respectively, does not rule out a strong possibility of DnaA and cisII or cisMP independent regulation of replication in these replicons and would be worth investigating independently.
Materials and Methods
Bacterial strains, plasmids, and materials
D. radiodurans R1 (ATCC13939) was grown in TGY (1% tryptone, 0.1% glucose, and 0.5% yeast extract) medium at 32°C. E. coli strain NovaBlue was used for cloning and maintenance of all the plasmids, whereas E. coli strain BL21 (DE3) pLysS was used for the expression of recombinant proteins. The pNOKOUT plasmid (a non-replicative plasmid in D. radiodurans) was used for generating gene knockouts and bioassay of cis-elements for origin of replication function in D. radiodurans. Standard protocols for all the recombinant techniques were used as described earlier (Green & Sambrook, 2012). Molecular biology grade chemicals and enzymes were procured from Merck Inc. and New England Biolabs. Radiolabeled nucleotides were purchased from the Board of Radiation and Isotope Technology, Department of Atomic Energy, India (BRIT). All the oligonucleotides used in this study are listed in Table S1 and all bacterial strains and plasmids are listed in Table S2.
Bioinformatic analysis
The nucleotide sequences close to the origin region and upstream to parAB operons in the circular map of ChrII (99–544 bp region) and MP (177,446–417 bp region) were analysed for potential repeats and consensus sequence using Mellina II web tool (Okumura et al, 2007) and WebLogo online tool (Crooks et al, 2004) and tentatively named as cisII and cisMP, respectively. The putative DnaA boxes in cisII were predicted using DOriC database (DoriC accession number–ORI10030006) (Luo & Gao, 2019). The DnaA boxes in cisMP were marked based on sequence homology with known DnaA boxes from other bacteria.
Cloning of cis-elements and their maintenance
The cisII and cisMP were PCR-amplified from genomic DNA of D. radiodurans using cisIIFw/cisIIRw and cisMPFw/cisMPRw primers, respectively (Table S1). The cisII was cloned at XbaI site, whereas cisMP at ApaI and EcoRI sites in pNOKOUT (Khairnar et al, 2008) to yield pNOKcisII and pNOKcisMP plasmids, respectively (Table S2). These plasmids were transformed in both wild type and recA mutant of D. radiodurans and transformants were scored at kanamycin (6 μg/ml) containing TGY agar plates. For plasmid maintenance, the transformants were streaked on kanamycin containing TGY agar plates. The bacterial growth was also monitored in TGY liquid medium in the presence of antibiotic as required, at 32 °C and OD 600 nm was monitored using Synergy H1 Hybrid Multi-mode microplate reader, Bio-Tek. The maintenance of pNOKcisII and pNOKcisMP plasmids in D. radiodurans was confirmed with antibiotic resistance as well as by isolating these plasmids from transformant as described earlier (Meima & Lidstrom, 2000). The isolated plasmid DNA was digested with XbaI in the case of pNOKcisII and ApaI and EcoRI in the case of pNOKcisMP, and the restriction digestion pattern was compared with the original plasmid on the agarose gel.
Protein purification
The recombinant DnaA of D. radiodurans expressing on pETDnaA in E. coli BL21 (DE3) pLysS was purified by Ni-NTA affinity chromatography as described earlier (Maurya & Misra, 2020 Preprint). In brief, E. coli cells carrying pETDnaA were induced with 0.5 mM isopropyl-β-D-thio-galacto-pyranoside (IPTG) and cell pellet was suspended in buffer A (20 mM Tris–HCl, pH 7.6, 300 mM NaCl and 10% glycerol), 0.5 mg/ml lysozyme, 1 mM PMSF, 1 mM MgCl2, 0.05% NP-40, 0.05% Triton X-100, and protease inhibitor cocktail. The resultant mixture was sonicated on ice and cell lysate was centrifuged at 13,500g for 30 min at 4°C and the clear supernatant was purified through Ni-NTA column chromatography (GE Healthcare). The recombinant ParBs encoded on chromosome II (ParB2) and megaplasmid (ParB3) in D. radiodurans were purified from recombinant E. coli using Ni-NTA affinity chromatography (Maurya et al, 2019b). Fractions with >95% purity were pooled and purified from HiTrap Heparin HP affinity columns (GE Healthcare Life Sciences) using a linear gradient of 20–300 mM NaCl, followed by precipitation with 30% wt/vol ammonium sulphate at 8°C, and molecular exclusion chromatography using the standard protocol. Histidine-tag was removed from the recombinant proteins by digesting with Factor Xa protease followed by passing through Ni affinity column. The absence of his-tag in the protein eluted in flow-through fraction was confirmed using poly histidine antibodies and the concentration was determined by taking OD at 280 nm in NanoDrop (Synergy H1 Hybrid Multi-Mode Reader Biotek).
DNA–protein interaction studies
DNA binding activity of DnaA, ParB2, and ParB3 was monitored using electrophoretic mobility shift assay as described earlier (Charaka & Misra, 2012; Maurya et al, 2019a) with different versions of cis-element as DNA substrates. In brief, the full-length cisII and cisMP and their repeat derivatives (Fig S4A and B) were PCR-amplified using sequence-specific primers (Table S1) except the two repeats and one repeat that were chemically synthesized and annealed with the complementary strand. The [32P] labeled DNA probes were used for DNA binding assay. For the competition assay, saturating concentrations of DnaA or ParBs with radiolabelled DNA substrates were chased with different concentrations of unlabeled non-specific DNA (NS-DNA) of similar length (Table S1) and a 10-fold higher concentration of same unlabeled cis sequence. The products were resolved on 6–10% native PAGE gel as required. The DNA band intensity was determined densitometrically using Image J software, and the fractions of bound DNA were plotted against different protein concentrations using GraphPad Prism 5. The Kd value for the curve fitting of individual plots was determined.

ts. (A, B) Schematic representation of cisII (A) and cisMP (B) elements and their repeat variants PCR amplified and used for DNA–protein interaction studies.
Construction of deletion mutants and cell survival studies
The constructs for generating knockout of both cis-elements were made in pNOKOUT plasmid as described earlier (Khairnar et al, 2008). In brief, ∼500 bps upstream and ∼500 bps downstream fragments to the cisII and cisMP were PCR amplified with the primers (Table S1) and cloned at KpnI/EcoRI and BamHI/XbaI sites in pNOKOUT plasmid to yield pNOKCII and pNOKCM plasmids, respectively (Table S2). These plasmids were linearized with XmnI and transformed into D. radiodurans cells. The homozygous replacement of cisII and cisMP by nptII was achieved through several rounds of sub-culturing and then ascertained by diagnostic PCR using flanking primers (Table S1 and Fig S2A–C). The confirmed mutant cells were maintained in the absence of selection pressure but ensured for antibiotic resistance before the experiments.
The survival of ΔcisII and ΔcisMP deletion mutants was monitored under normal conditions and in response to different doses of γ radiation as described earlier (Misra et al, 2006). In brief, the cells were grown in the absence and presence of antibiotics. These cells were plated on TGY agar plate with and without antibiotics under normal conditions. These cells were independently exposed to different doses of γ-radiation (2, 4, 6, and 9 kGy) at the dose rate of 1.81 kGy/h using Gamma Cell 5000 irradiator (60Co; Board of Radiation and Isotopes Technology, DAE). The equal number of irradiated cells and their SHAM controls were washed in PBS and the different dilutions were plated in replicates on TGY agar plates with antibiotics, as required. The plates were incubated at 32°C for 45–48 h and CFUs were counted.
Estimation of genome copy number
The copy number of genomic replicon and mini replicons like pNOKcisII and pNOKcisMP was determined in the exponentially growing D. radiodurans cells using modified protocols of Breuert et al (2006) as described earlier in Maurya et al (2019a). In brief, the density and cell numbers of the independent sample were normalized for a fixed OD 600 nm and by CFUs. These cells lysed in buffer containing 10 mM Tris, pH 7.6, 1 mM EDTA, and 4 mg/ml lysozyme followed by repeated cycles of freezing and thawing. The clear cell-free extract was used for quantitative Real-Time PCR. The qPCR analysis involves three different genes of similar PCR efficiency from each genomic replicon of D. radiodurans. For instance, ftsE, ftsK, and ftsZ for chromosome I; DR_A0155, DR_A0182, and pprA for chromosome II; DR_B0003, DR_B0030, and DR_B0104 for megaplasmid and DR_C0001, DR_C0018, and DR_C0034 for small plasmid (Table S1). For mini replicons, the nptII (KanR) and bla (AmpR) genes of plasmid backbones (Table S1) were used for copy number determination. The 300 bp PCR amplified DNA fragments were quantified by nanodrop and DNA concentrations were calculated using the molecular mass computed with “oligo calc” (www.basic.northwestern.edu/biotools). A serial dilution was made for each standard fragment and qPCR was carried out by following the MIQE guidelines using Roche Light cycler (Bustin et al, 2009) and the cycle threshold (Ct) values were determined. The copy number of each replicon is quantified by comparing the results with a standard plot. The average copy number reflected from all three genes per replicon was analysed using statistical analysis of the data obtained from the three independent biologic replicates for each sample.
Construction of tetO insertion derivatives of the genomic replicons in D. radiodurans
The 240 repeats of tetO operator cassette from pLAU44 plasmid (Lau et al, 2003) were integrated near the putative “origin of replication” at 1.5° in ChrI, 4° in ChrII, and 4.5° in MP of the circular map of these replicons in D. radiodurans. For that, the PCR amplifications of 10,713–11,715 bp region corresponding to 1.5° of ChrI using ChrI(1.5°)Fw and ChrI(1.5°)Rw primers, 4659–c5691 bp region corresponding to 4° of ChrII using ChrII(4°)Fw and ChrII(4°)Rw primers, and 2,203–3,000 bp region corresponding to 4.5° of MP using Mp (4.5°)Fw and Mp(4.5°)Rw were carried out. These PCR fragments were cloned in pLAU44 at XbaI-ScaI sites to yield p44Ch1, p44Ch2 and p44MP plasmids having homologous sequences of ChrI, ChrII, and MP, respectively. Furthermore, an expressing cassette of spectinomycin resistance gene was amplified from pVHS559 (Charaka & Misra, 2012) using SpecFw and SpecRw primers and cloned at NheI-XhoI sites in p44Ch1, p44Ch2, and p44MP to give p44SCh1, p44SCh2, and p44SMP, respectively. These constructs were confirmed by restriction digestion, PCR and DNA sequencing. The p44SCh1, p44SCh2, and p44SMP plasmids were separately transformed into wild type, and ΔcisII and ΔcisMP mutants of D. radiodurans. The transformants were scored under antibiotics selection and the homogenous insertion of tetO repeats in the genome was achieved through several rounds of subculturing. This was ascertained by diagnostic PCR using different sets of primers (Table S1). The resultant strains of D. radiodurans were named with suffix as R1::ChrI-tetO, R1::ChrII-tetO, R1::MP-tetO, ΔcisII::ChrI-tetO, ΔcisII::ChrII-tetO, ΔcisII::MP-tetO, ΔcisMP::ChrI-tetO, ΔcisMP::ChrII-tetO, and ΔcisMP::MP-tetO as listed in Table S2.
Construction of TetR-GFP expression plasmid in D. radiodurans
The coding sequence of tetR was PCR amplified from pLAU53 (Lau et al, 2003) using gene-specific primers (Table S1) and cloned at SacI and SalI sites in pDSW208 (Weiss et al, 1999) to give pDTRGFP plasmid. Furthermore, tetR-gfp was PCR amplified from pDTRGFP plasmid using TetRscIApIFw and GFPXbaIRw primers and cloned in pRADgro (Khairnar et al, 2008) at ApaI-XbaI sites to produce pRADTRGFP (Table S2). The constitutive expression of TetR-GFP from pRADTRGFP plasmid in D. radiodurans was confirmed by fluorescence microscopy and the Western blotting using anti-GFP antibodies (Fig S5A and B). The tetO operator containing cells of D. radiodurans R1, ΔcisII and ΔcisMP mutants (Table S2) expressing TetR-GFP on pRADTRGFP plasmid were observed under a fluorescence microscope as described earlier (Maurya et al, 2019a). In brief, overnight (14 h) grown cultures were harvested, washed with PBS and stained with DAPI (4′,6-diamidino-2-phenylindole, dihydrochloride; 0.2 mg/ml) for nucleoid and Nile red (1 mg/ml) for membrane. These samples were imaged through differential interference contrast, DAPI, FITC, and TRITC channels under an inverted fluorescence microscope (Olympus IX83) fitted with an Olympus DP80 CCD monochrome camera. The images were aligned and processed using in-built software “CellSens.” A population of ∼200 cells was analysed for results and data were presented as a scattered plot. One representative image belonging to more than 90% population is presented separately as differential interference contrast, DAPI, Nile Red, and GFP and merged of all these. Experiments were repeated independently to ensure the reproducibility and significance of these data.
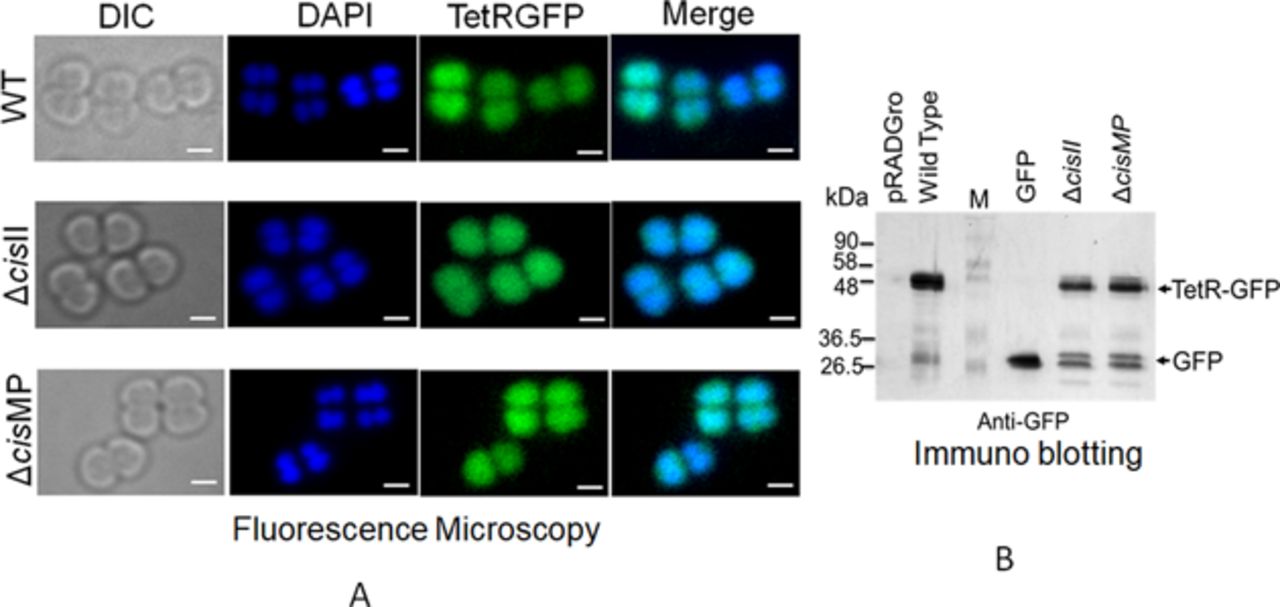
(A, B) The expression of TetR-GFP in both wild type and cis mutants of D. radiodurans was confirmed microscopically (A) and through immunoblotting (B). The TetR-GFP expression in the absence of tetO element was found to be uniformly spread in the cell.
Data Availability
This article contains data that have been recorded using low-throughput technologies and are available anytime for cross references.
Acknowledgements
The authors are grateful to Dr SK Apte for his comment on the work and results, and Dr Swathi Kota, Dr YS Rajpurohit, and Dr Sheetal Uppal for their technical inputs. The authors also thank Ms Shruti Mishra and Ms Reema Chaudhary for reading through the manuscript, their comments, and help in fluorescence imaging. GK Maurya is grateful to the Department of Atomic Energy, India, for the research fellowship.
Author Contributions
GK Maurya: conceptualization, resources, data curation, formal analysis, investigation, methodology, and writing—original draft, review, and editing.
HS Misra: conceptualization, resources, data curation, formal analysis, supervision, funding acquisition, validation, investigation, visualization, methodology, project administration, and writing—original draft, review, and editing.
Conflict of Interest Statement
The authors declare that they have no conflict of interest.
- Received July 22, 2020.
- Revision received November 4, 2020.
- Accepted November 4, 2020.
- © 2020 Maurya et al.
This article is available under a Creative Commons License (Attribution 4.0 International, as described at https://creativecommons.org/licenses/by/4.0/).
References




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- No citing articles found.