



Article Figures & Data
Figures
- Figure 1. Profilin is present in centrosomes throughout the cell cycle.
Fluorescence microscopy revealed the co-localization of profilin and γ-tubulin. (A) Ab staining of profilin and γ-Tb in B16 cells after a brief pretreatment with 0.1% Triton X-100 before fixation (see text). High-resolution confocal microscopy (AiryScan). (B) B16 cells expressing citrine-profilin (a, d, g; green) were captured at different cell cycle stages. Cells were permeabilized with 10 μM digitonin, fixed, and stained for γ-tubulin (b, e, h; red). Superposition of images in (c, f, i). The cell in (d, e, f) was captured at an early G2-stage of the cell cycle and the one in (g, h, i) at late G2/prophase. Micrographs were obtained by spinning disk microscopy. Scale bars (A) and (B): 10 μm.
- Figure S1. Profilin accumulates in centrosomes during mitosis.
Immunofluorescence staining of profilin and γ-tubulin in B16 cells followed by intensity measurements of centrosomal fluorescence reveals increased labeling of Pfn in mitotic cells. (A) Cells captured during interphase (top) and during metaphase (bottom). Wide-field microscopy after simultaneous staining for Pfn, γ-tubulin (γ-Tb) and DNA (DAPI). The centrosomal region is shown at higher magnification at the bottom left corner (inset); the arrow points to the centrosome. (B) Bar graph illustrating fluorescence intensity of Pfn labeling in a standardized circular area (1.2 μm2) around the centrosomes in interphase (n = 31) and metaphase cells (n = 44) as determined by wide-field microscopy. Mean ± SEM, **P < 0.01. (C) High-resolution confocal microscopy (AiryScan) of a metaphase cell after simultaneous staining for Pfn, α-tubulin (α-Tb), and DNA (DAPI). Scale bars 10 μm.
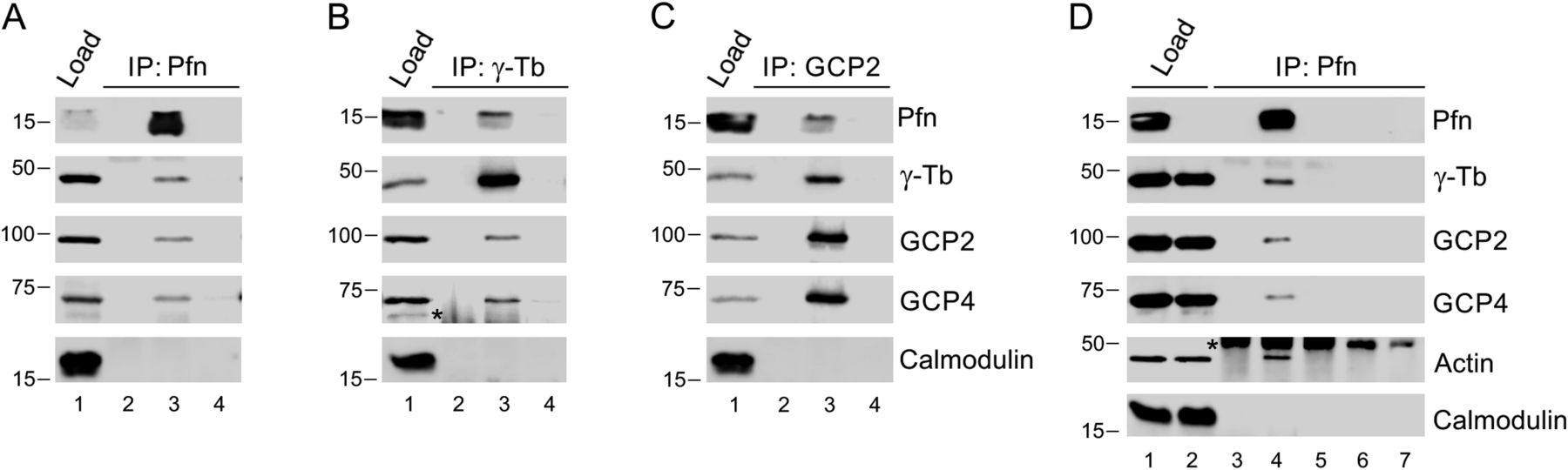
- Figure 2. Profilin interacts with γ-TuRC.
(A, B, C) Extracts of B16 cells were precipitated with immobilized Abs to profilin (A), γ-tubulin (B) or GCP2 (C), followed by Western blotting and probing with Abs against profilin (Pfn), γ-tubulin (γ-Tb), GCP2, GCP4, and calmodulin (negative control) as indicated (C). In each blot (A, B, C), the sample lanes denoted 1–4 show: total cell extract, gel load (1); no cell extract (2); precipitated proteins (3); and Ab-free carrier (4). Note reciprocal precipitation in all three cases. (D) Extracts of B16 control (1, 4, 6) and profilin knock-out KO27 (2, 5, 7) cells were precipitated with immobilized Abs to profilin, followed by Western blotting and probing with Abs against profilin (Pfn), γ-tubulin (γ-Tb), GCP2, GCP4, actin, and calmodulin (negative control) as indicated. The sample lanes denoted 1–7 show: total cell extract, gel load (1, 2); no cell extract (3); precipitated proteins (4–5); and Ab-free carrier (6–7). Note co-precipitation of γ-tubulin, GCP2, GCP4, and actin with profilin Ab occurs in B16 cells solely. (B, D) Asterisks (*) in panels (B) and (D), lanes 2 and 3, and 3–5, respectively, denote cut off bands reflecting precipitating Abs.
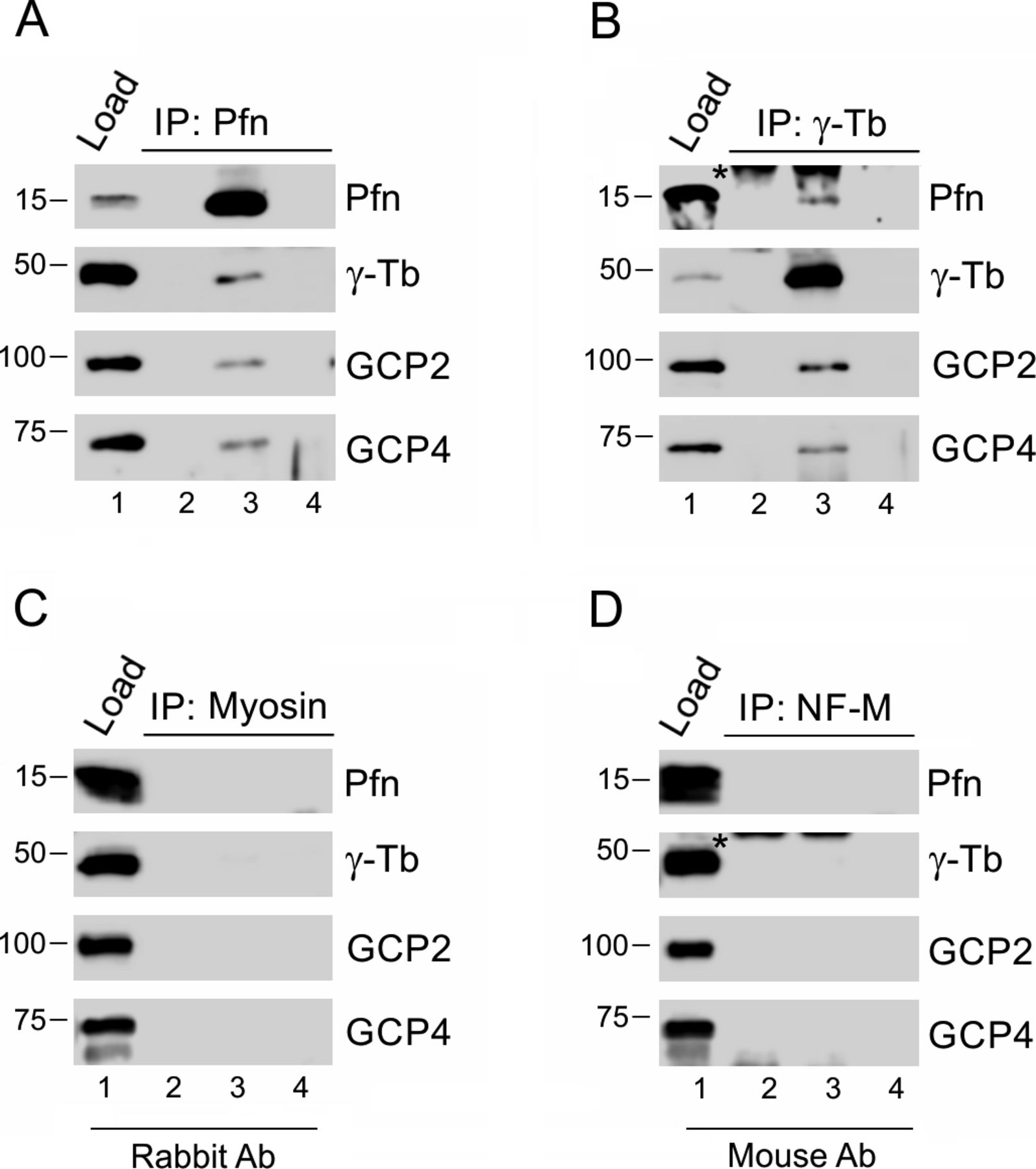
- Figure S2. Profilin associates with γ-tubulin complex proteins in Caco-2 cells, and isotype controls of immunoprecipitations are negative.
(A, B) Extracts from Caco-2 cells were precipitated with immobilized Abs to profilin (A) and γ-tubulin (B), respectively, followed by Western blot analysis, using Abs against profilin (Pfn), γ-tubulin (γ-Tb), GCP2 and GCP4. Sample lanes 1–4 represent: total cell extract, gel load (1); no cell extract (2); precipitated proteins (3); carrier without Ab (4). (C, D) extracts of B16 cells were incubated with immobilized isotype control Abs, using rabbit Ab to myosin (C) and mouse mAbs (IgG2a) to neurofilament NF-M (D), respectively. (A, B) Subsequent Western blotting and analyzes were performed as in (A, B), using the same sample denotations. (B, D) Asterisks (*) in panels (B) and (D), lanes 2 and 3 denote cut off bands reflecting precipitating Ab.
- Figure 3. Profilin modulates microtubule regrowth.
(A) B16 control and profilin knock-out (KO27) cells visualized after nocodazole washout and 2 min of recovery in drug-free medium at 37°C followed by processing for fluorescence labeling of microtubules (α-tubulin; α-Tb) and centrosomes (γ-tubulin; γ-Tb). Note the more prominent array of nascent microtubules extending from the centrosome in the KO27 cells compared with the control. Scale bar: 10 μm. (B) Box plots illustrating the distribution of α-tubulin fluorescence intensity (arbitrary units, AU) determined 3 min after drug washout at 28°C within a 1.0-μm region of interest in B16 control and KO27 cells, respectively; n = 330 (control; Ctr) and 322 (KO27; Pfn KO). (C) As in (B), but γ-tubulin fluorescence intensity; n = 339 (control) and 284 (KO27). The data in (B, C) are based on three independent experiments and >90 cells for each experimental condition. The thick and thin lines within each box represent the mean and median (50th percentile), respectively, whereas bottom and top represent the 25th and 75th percentiles. The whiskers below and above the box indicate the 10th and 90th percentiles; ****P < 0.0001. (D) Microtubule nucleation rate in KO27 cells relative to control cells. Three independent experiments with at least 13 cells counted in each experiment, n = 39 (control) and 55 (KO27). The thick and thin lines within the dot box plots represent mean ± SD; ****P < 0.0001. (E) Time-lapse imaging of control and KO27 cells expressing EB3-mNeonGreen. Tracks of EB3 comets captured during 10 s are shown. Scale bar: 10 μm.
- Figure S3. Nocodazole-resistant microtubules and microtubule organization in profilin knock-out cells (KO27).
(A) Nocodazole-resistant microtubules in B16 control (Ctr) and KO27 (Pfn KO) cells simultaneously stained by Abs to α-tubulin (α-Tb) and γ-tubulin (γ-Tb). Fixation F/Tx/M (see the Materials and Methods section). Scale bar, 10 μm. (B) Microtubule organization in B16 control (Ctr) and KO27 (Pfn KO) cells stained by Ab to α-tubulin (α-Tb). Arrowheads in left panel (Ctr) denote the tip of the cell edge. Scale bar, 10 μm. (C) Bar graph showing increased α-tubulin fluorescence in two randomly picked areas of standardized size (144 μm2) in the lamellae of control (n = 29) and KO27 (n = 33) cells, respectively. Mean ± SEM, ***P < 0.001.
- Figure 4. Phenotypic rescue of increased microtubule nucleation in KO27 cells.
(A) Immunoblot analysis of whole-cell lysates of B16 control cells expressing citrine-cathepsin B (Ctr+Citrine-Cath), profilin knock-out KO27 cells expressing citrine-cathepsin B (Pfn KO+Citrine-Cath), and KO27 cells rescued by citrine-profilin (Pfn KO+Citrine-Pfn). Blots were probed with Abs to GFP to detect citrine-tagged proteins, to EB1 to detect co-expressed EB1-tdTomato, profilin (Pfn), and to actin (loading control). (B) Comparison of the microtubule nucleation rate observed in KO27 (Pfn KO) cells expressing Citrine-cathepsin B and Citrine-profilin, respectively, and in control cells (Ctr) expressing Citrine-cathepsin B. Three independent experiments and at least 10 cells analyzed in each experiment, n = 36 (Ctr+Citrine-Cath), n = 31 (Pfn KO+Citrine-Cath), and n = 30 (Pfn KO+Citrine-profilin); abbreviations as in left panel. The thick and thin lines within dot box plots represent mean ± SD; ****P < 0.0001.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
