




Article Figures & Data
Figures
- Figure 1. Cleavage of coronavirus S protein.
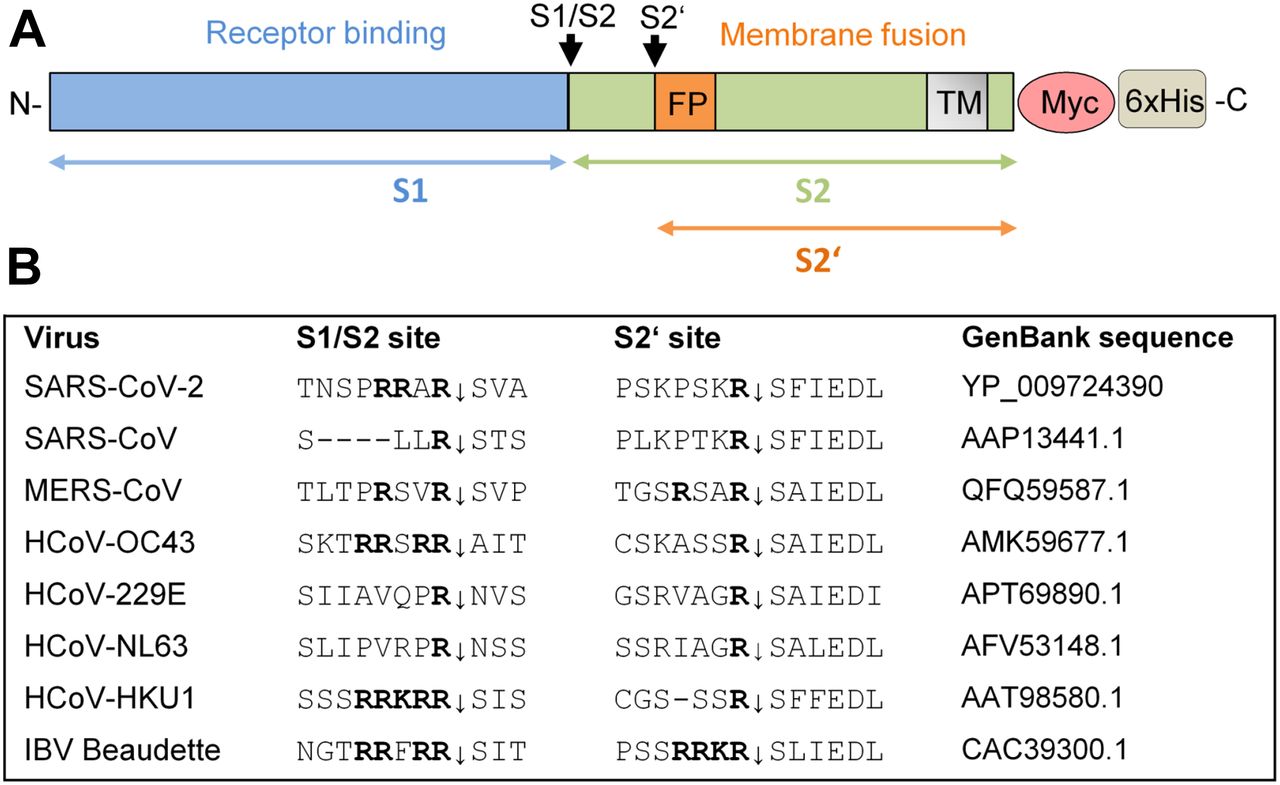
(A) Schematic representation of the SARS-CoV-2 precursor and the S1 and S2 subunits. Fusion peptide (FP), and transmembrane domain (TM) are indicated. The S1/S2 and S2′ cleavage sites and subunits S1, S2, and S2′ are indicated by black and colored arrows, respectively. For immunochemical detection, recombinant S is expressed with a C-terminally fused Myc-6xHis-tag peptide in our study. (B) Alignment of the amino acid sequences at the S1/S2 and S2′ cleavage site of the S proteins of different human coronaviruses (HCoV) and avian infectious bronchitis virus strain Beaudette.
- Figure 2. Cleavage of SARS-CoV-2 S by furin and TMPRSS2.
(A) Fluorescence resonance energy transfer substrates of the S protein S1/S2 sites of the indicated CoVs. M1 and M2 are mutants of the SARS-CoV-2 S1/S2 site with substitution of A → K or A → R in P2 position. IBV, avian infectious bronchitis virus strain Beaudette. Cleavage by furin is indicated in red. (B) Cleavage of the fluorescence resonance energy transfer substrates (20 μM) by furin (0.5 nM). Cleavage efficiency of SARS-CoV-2_M2 was set as 100%. (C) Cleavage of SARS-CoV-2 S by furin and TMPRSS2 in HEK293 cells. Cells were co-transfected with pCAGGS-S-Myc-6xHis and either empty vector or pCAGGS-TMPRSS2. Cells were then incubated in the absence or presence of aprotinin or furin inhibitor MI-1851 (50 μM each) for 48 h. Cell lysates were subjected to SDS–PAGE and Western blot analysis using antibodies against the C-terminal Myc-tag. For each Western blot lanes are spliced together from one immunoblot from one experiment. β-actin was used as loading control.
Source data are available for this figure.
Source Data for Figure 2[LSA-2020-00786_SdataF2_F3.pptx]
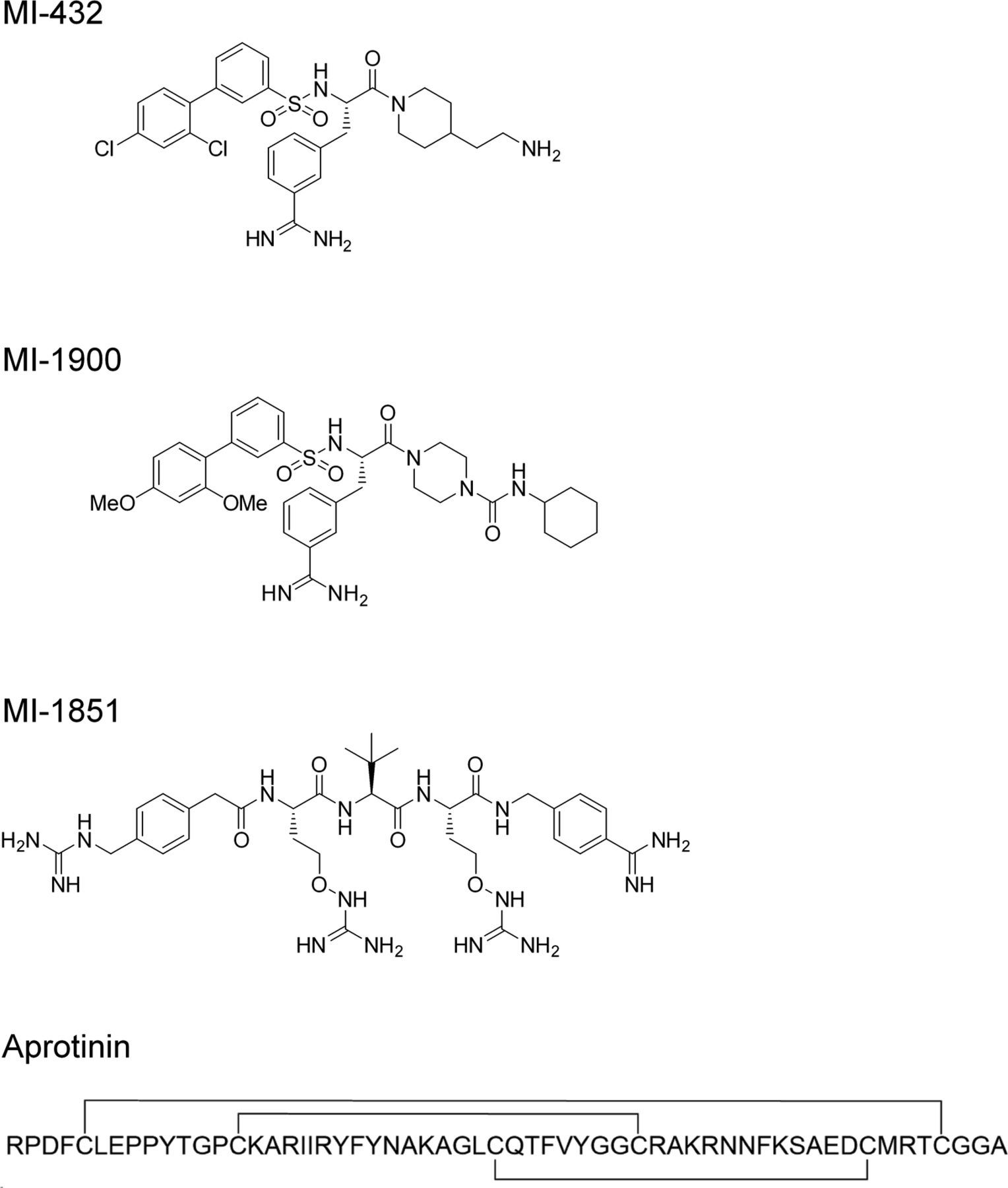
- Figure S1. Structural formulas of peptide mimetic inhibitors MI-432, MI-1900 and MI-1851 and the linear amino acid sequence of bovine aprotinin (24).
Aprotinin contains three disulfide bonds (indicated by lines).
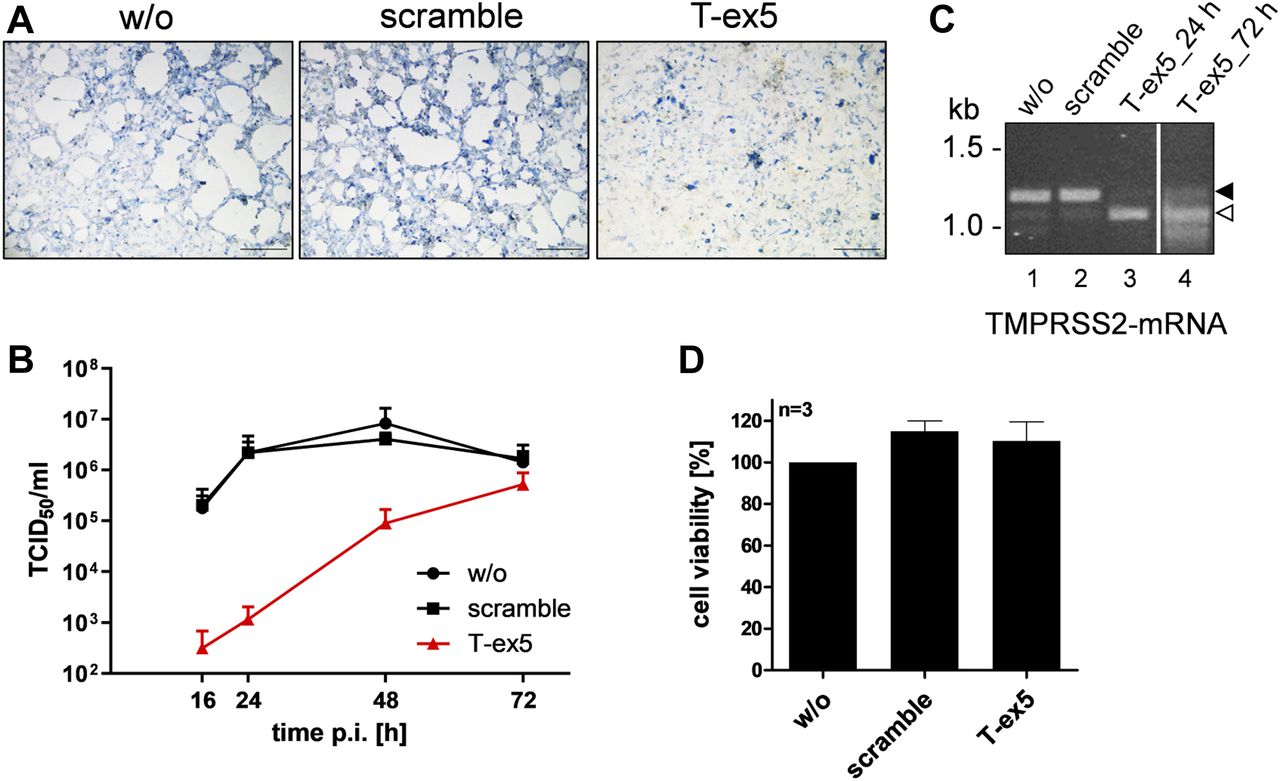
- Figure 3. Knockdown of TMPRSS2 expression by PPMO T-ex5 inhibits multicycle replication of SARS-CoV-2 in Calu-3 cells.
(A) Multicycle replication of SARS-CoV-2 in T-ex5–treated Calu-3 cells. Cells were treated with 25 μM T-ex5 or control PPMO (scramble) for 24 h or remained without treatment (w/o). Cells were then inoculated with SARS-CoV-2 at a MOI of 0.001 for 1 h 30 min, the inoculum was removed and the cells further incubated in the absence of PPMO for 72 h. Cells were fixed and immunostained using a serum against SARS-CoV. Virus-positive cells are stained in blue. Scale bars indicate 500 μm. (B) Calu-3 cells were treated with PPMO for 24 h and then infected with SARS-CoV-2 for 72 h as described above. Virus titers in supernatants were determined by tissue culture infection dose 50% (TCID50) end point dilution at indicated time points. Results are mean values ± SD of three independent experiments. (C) Analysis of TMPRSS2-mRNA in PPMO-treated Calu-3 cells. Cells were treated with 25 μM T-ex5, scramble PPMO or remained untreated (w/o) for 24 h (lanes 1–4). T-ex5–treated cells were inoculated with SARS-CoV-2 as described above and incubated in the absence of PPMO for 72 h (lane 4). Total RNA was isolated and analyzed by RT-PCR using primers designed to amplify 1,228 nt of full-length TMPRSS2-mRNA. Full-length and truncated PCR products lacking exon 5 are indicated by filled and open arrow heads, respectively. (D) Effect of PPMO treatment on Calu-3 cell viability. Calu-3 cells were treated with scramble or T-ex5 PPMO (25 μM) for 24 h. Cell viability of untreated (w/o) cells was set as 100%. Results are mean values ± SD (n = 3).
Source data are available for this figure.
Source Data for Figure 3[LSA-2020-00786_SdataF2_F3.pptx]
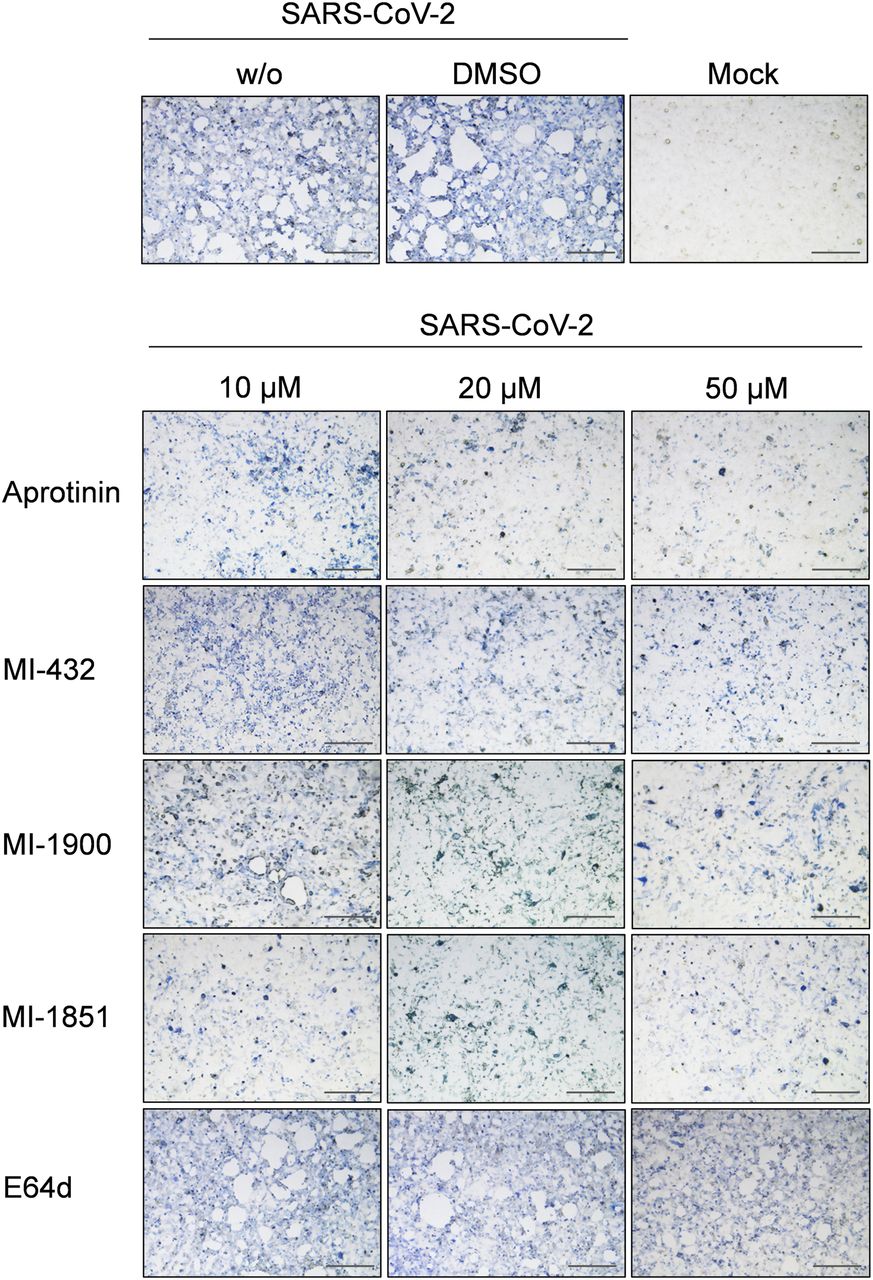
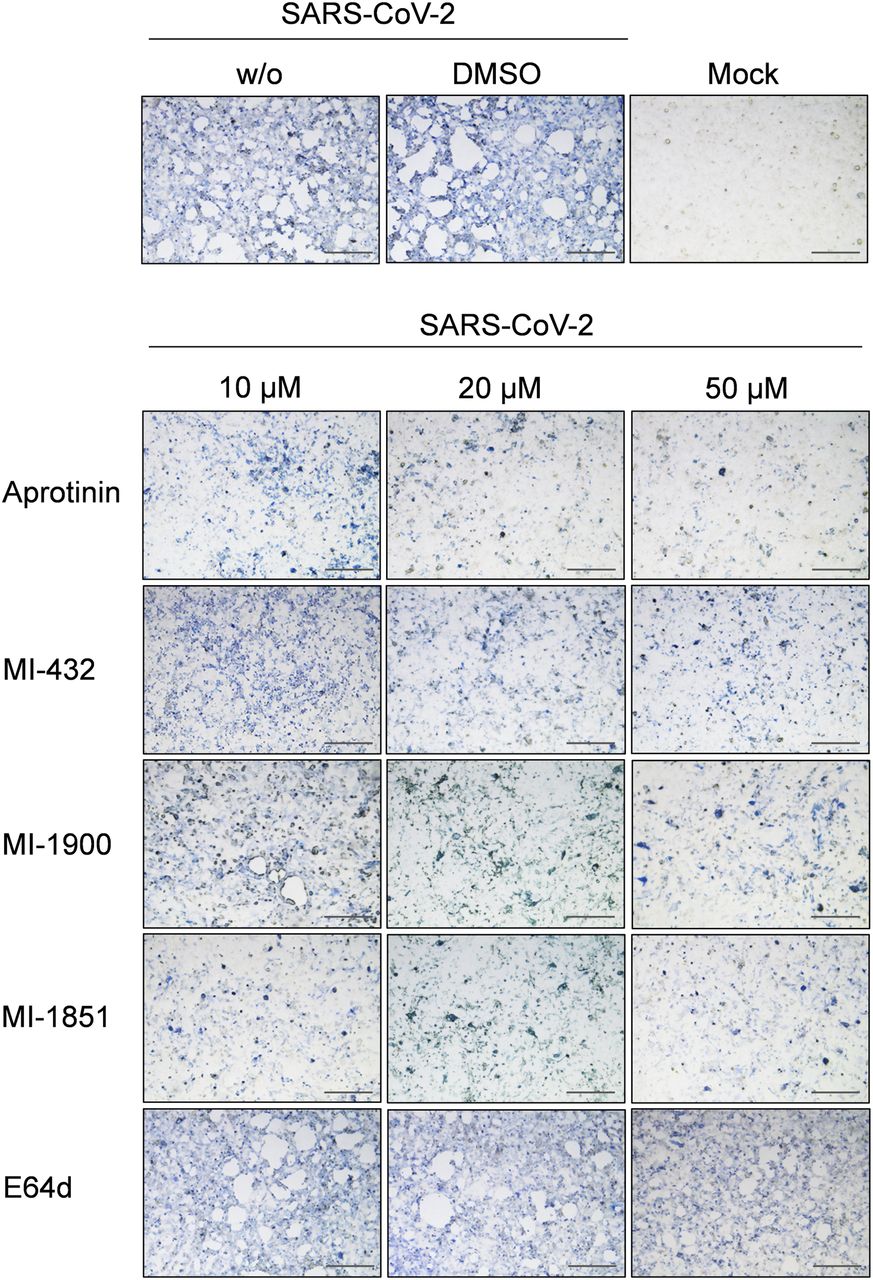
- Figure 4. Inhibition of SARS-CoV-2 multiplication in human airway cells by inhibitors of furin and TMPRSS2.
Calu-3 cells were inoculated with SARS-CoV-2 at a low MOI of 0.001 and then incubated in the presence of inhibitors of TMPRSS2 (aprotinin, MI-432, and MI-1900), furin (MI-1851), and endosomal cathepsins (E64d), respectively, at the indicated concentrations. Cells were fixed and immunostained using a rabbit serum against SARS-CoV at 72 h p.i. Virus-positive cells are stained in blue or dark gray depending on the staining intensity. Cells infected in the absence of inhibitors (w/o), in the presence of DMSO (0.5%) and noninfected cells (mock) were used as controls. Scale bars indicate 500 μm. Images are representatives of three independent experiments.
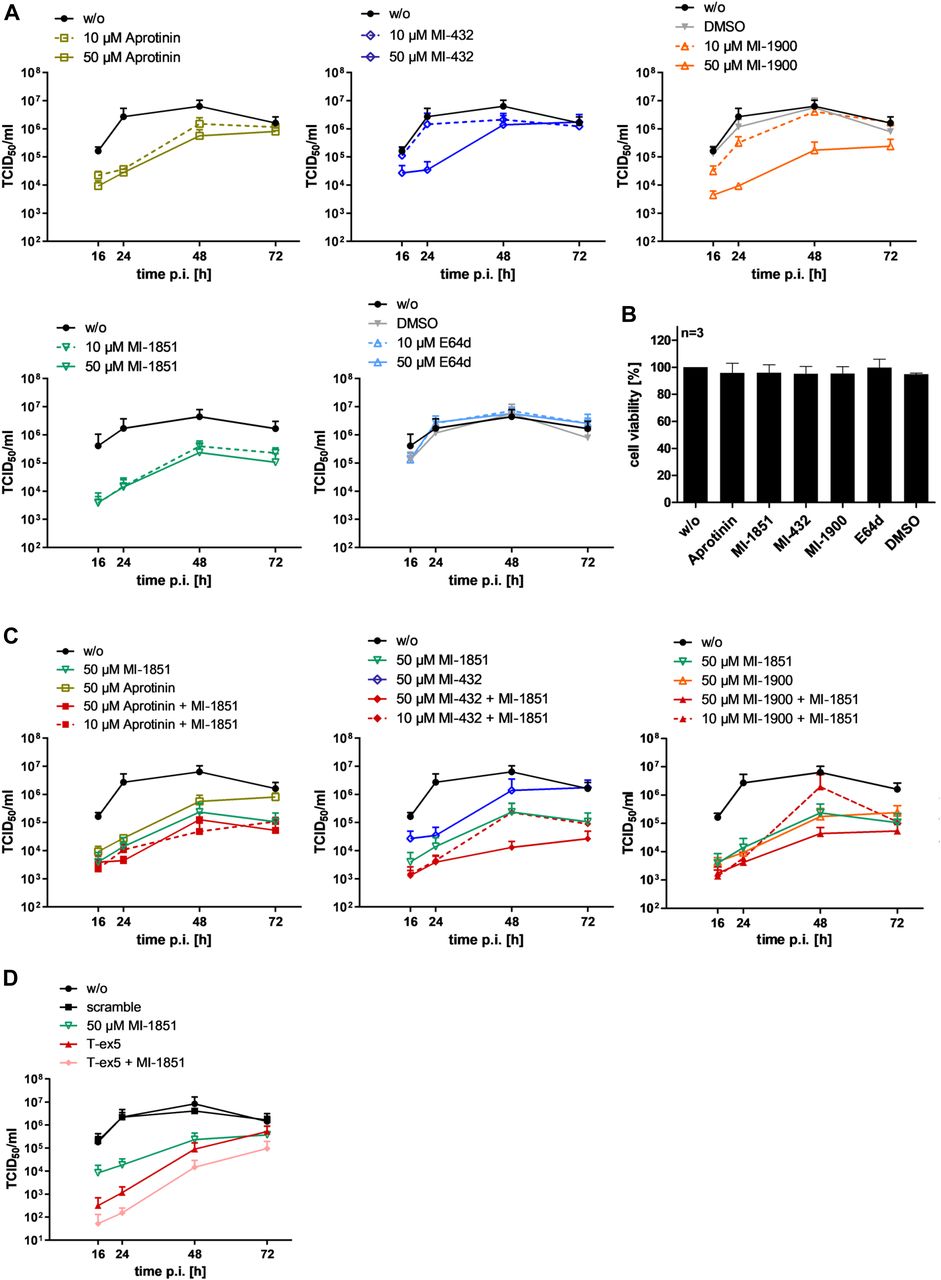
- Figure 5. Inhibition of SARS-CoV-2 multicycle replication in human airway epithelial cells by inhibitors of TMPRSS2 and furin.
(A) Calu-3 cells were inoculated with SARS-CoV-2 at a low MOI of 0.001 and then incubated in the absence (w/o) or presence of inhibitors of TMPRSS2 (aprotinin, MI-432, and MI-1900), furin (MI-1851), and endosomal cathepsins (E64d), respectively, or DMSO (0.5%), at the indicated concentrations. At 16, 24, 48, and 72 h postinfection (p.i.), supernatants were collected, and virus replication was determined by tissue culture infection dose 50% (TCID50) titration at indicated time points. Data are mean values ± SD of three to five independent experiments. (B) Effect of inhibitor treatment on cell viability. Calu-3 cells were treated with the indicated protease inhibitor (50 μM) for 72 h. Untreated cells (w/o) and DMSO treated cells were used as controls. Cell viability of untreated cells was set as 100%. Results are mean values ± SD (n = 3). (C) Antiviral activity of combinations of TMPRSS2 and furin inhibitors against SARS-CoV-2 in human airway epithelial cells. Calu-3 cells were inoculated with SARS-CoV-2 at an MOI of 0.001 as described above and then incubated in the presence of single protease inhibitors or inhibitor combinations at the indicated concentrations. Virus titers in supernatants were determined by TCID50 at 16, 24, 48, and 72 h p.i. Data are mean values ± SD of three independent experiments. (D) Calu-3 cells were treated with PPMO for 24 h, then infected with SARS-CoV-2 as described above and incubated in the absence of PPMO (w/o, scramble and T-ex5) and with or without 50 μM of furin inhibitor treatment (MI-1851) for 72 h. At 16, 24, 48, and 72 h p.i., supernatants were collected, and viral titers were determined by TCID50 at indicated time points. Data are mean values ± SD (n = 2).
- Figure S2. Cleavage analysis of SARS-CoV-2 S2′ site by furin.
(A) Fluorescence resonance energy transfer substrates of the S protein S2′ sites of the indicated CoVs. M3 is a mutant of the SARS-CoV-2 S2′ site with substitution of P → R in P4 position. IBV, avian infectious bronchitis virus strain Beaudette. Cleavage by furin is indicated in red. (B) Cleavage of the fluorescence resonance energy transfer substrates (20 μM) by furin (0.5 nM). Cleavage efficiency of IBV Beaudette was set as 100%.
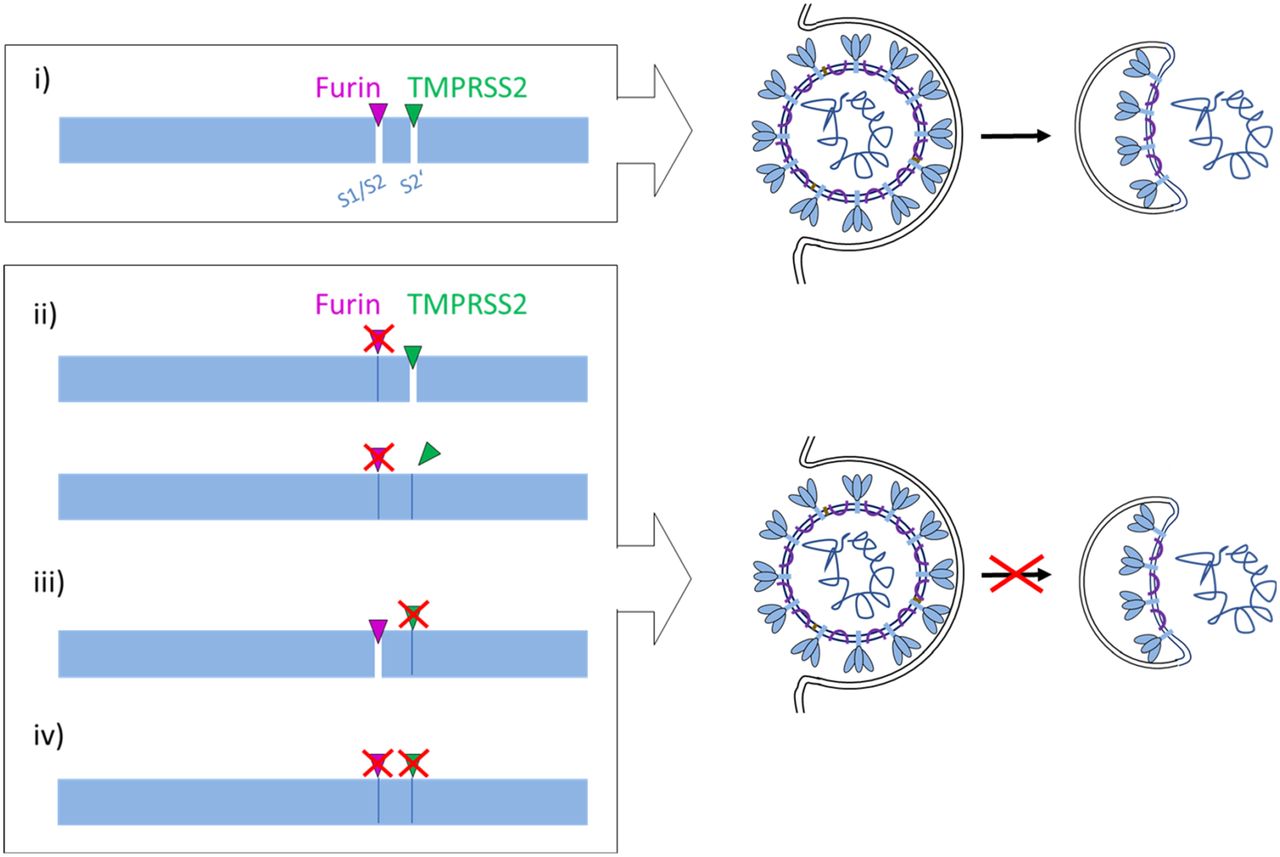
- Figure 6. Proposed processing of SARS-CoV-2 spike protein S by TMPRSS2 and furin.
(i) S must be cleaved at two sites, S1/S2 and S2′, to trigger fusion of viral and cellular membranes during virus entry to release the virus genome into the host cell. CoV S cleavage is believed to occur sequentially, with cleavage at the S1/S2 site occurring first and subsequent cleavage at the S2′ site. Furin processes the S1/S2 site, whereas TMPRSS2 cleaves at the S2′ site, and both proteases cannot compensate each other. Inhibition of either furin (ii) or TMPRSS2 (iii) or simultaneous inhibition of both proteases (iv) renders the S protein fusion-inactive and prevents virus entry. Inhibition of TMPRSS2 prevents exposure of the fusion peptide at the N-terminus of the S2′ subunit (iii, iv). Inhibition of furin cleavage at the S1/S2 site may directly interfere with virus entry and membrane fusion by steric blockage of conformational changes (ii, upper scheme) or may prevent exposure of the S2′ site to TMPRSS2 (ii, lower scheme).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Related Articles
Cited By...
- In-silico docking platform with serine protease inhibitor (SERPIN) structures identifies host cysteine protease targets with significance for SARS-CoV-2
- Functional assessment of cell entry and receptor use for merbecoviruses
- Cholesterol and COVID-19--therapeutic opportunities at the host/virus interface during cell entry
- Efficacy of Host Cell Serine Protease Inhibitor MM3122 against SARS-CoV-2 for Treatment and Prevention of COVID-19
- TMPRSS2 activation of Omicron lineage Spike glycoproteins is regulated by TMPRSS2 cleavage of ACE2
- ACE2-EGFR-MAPK signaling contributes to SARS-CoV-2 infection
- Expression of TMPRSS2 is up-regulated by bacterial flagellin, LPS, and Pam3Cys in human airway cells
- Furin as a therapeutic target in cystic fibrosis airways disease
- Determinants and Mechanisms of the Low Fusogenicity and Endosomal Entry of Omicron Subvariants
- Rapalogs downmodulate intrinsic immunity and promote cell entry of SARS-CoV-2
- The diversity of the glycan shield of sarbecoviruses closely related to SARS-CoV-2
- Evolution of host protease interactions among SARS-CoV-2 variants of concern and related coronaviruses
- Tracking infectious entry routes of SARS-CoV-2
- ACE2-independent SARS-CoV-2 infection and mouse adaption emerge after passage in cells expressing human and mouse ACE2
- SARS-CoV-2 Omicron spike H655Y mutation is responsible for enhancement of the endosomal entry pathway and reduction of cell surface entry pathways
- A Suite of TMPRSS2 Assays for Screening Drug Repurposing Candidates as Potential Treatments of COVID-19
- Effect of SARS-CoV-2 spike mutations on its activation by TMPRSS2 and TMPRSS13
- IL4I1 binds to TMPRSS13 and competes with SARS-Cov2 Spike
- Metalloproteinase-dependent and TMPRSS2-independnt cell surface entry pathway of SARS-CoV-2 requires the furin-cleavage site and the S2 domain of spike protein
- A novel class of TMPRSS2 inhibitors potently block SARS-CoV-2 and MERS-CoV viral entry and protect human epithelial lung cells
- A diabetic milieu increases cellular susceptibility to SARS-CoV-2 infections in engineered human kidney organoids and diabetic patients
- Effect of natural mutations of SARS-CoV-2 on spike structure, conformation, and antigenicity
- A method for the generation of pseudotyped virus particles bearing SARS coronavirus spike protein in high yields
- FXa cleaves the SARS-CoV-2 spike protein and blocks cell entry to protect against infection with inferior effects in B.1.1.7 variant
- The PRRA Insert at the S1/S2 Site Modulates Cellular Tropism of SARS-CoV-2 and ACE2 Usage by the Closely Related Bat RaTG13
- The Furin-S2' Site in Avian Coronavirus Plays a Key Role in Central Nervous System Damage Progression
- On the Origin of SARS-CoV-2: Did Cell Culture Experiments Lead to Increased Virulence of the Progenitor Virus for Humans?
- SARS-CoV-2 Causes Severe Epithelial Inflammation and Barrier Dysfunction
- Large fluctuations of the fusion intermediate help SARS-CoV-2 capture host cell membranes
- Term Human Placental Trophoblasts Express SARS-CoV-2 Entry Factors ACE2, TMPRSS2, and Furin
- Crystal structure of inhibitor-bound human MSPL that can activate high pathogenic avian influenza
- Structural basis for GTP-induced dimerization and antiviral function of guanylate-binding proteins
- Coagulation factors directly cleave SARS-CoV-2 spike and enhance viral entry
- Crystal structure of inhibitor-bound human MSPL that can activate high pathogenic avian influenza
- SPINT2 controls SARS-CoV-2 viral infection and is associated to disease severity
- Spike Protein of SARS-CoV-2 Activates Macrophages and Contributes to Induction of Acute Lung Inflammations in Mice
- A monoclonal antibody against staphylococcal enterotoxin B superantigen inhibits SARS-CoV-2 entry in vitro
- The rocaglate CR-31-B (-) inhibits SARS-CoV-2 replication at non-cytotoxic, low nanomolar concentrations in vitro and ex vivo
- The SARS-CoV-2 and other human coronavirus spike proteins are fine-tuned towards temperature and proteases of the human airways
- Single-cell analyses reveal SARS-CoV-2 interference with intrinsic immune response in the human gut
- Safe and effective two-in-one replicon-and-VLP minispike vaccine for COVID-19
- A Tethered Ligand Assay to Probe the SARS-CoV-2 ACE2 Interaction under Constant Force