



Article Figures & Data
Figures
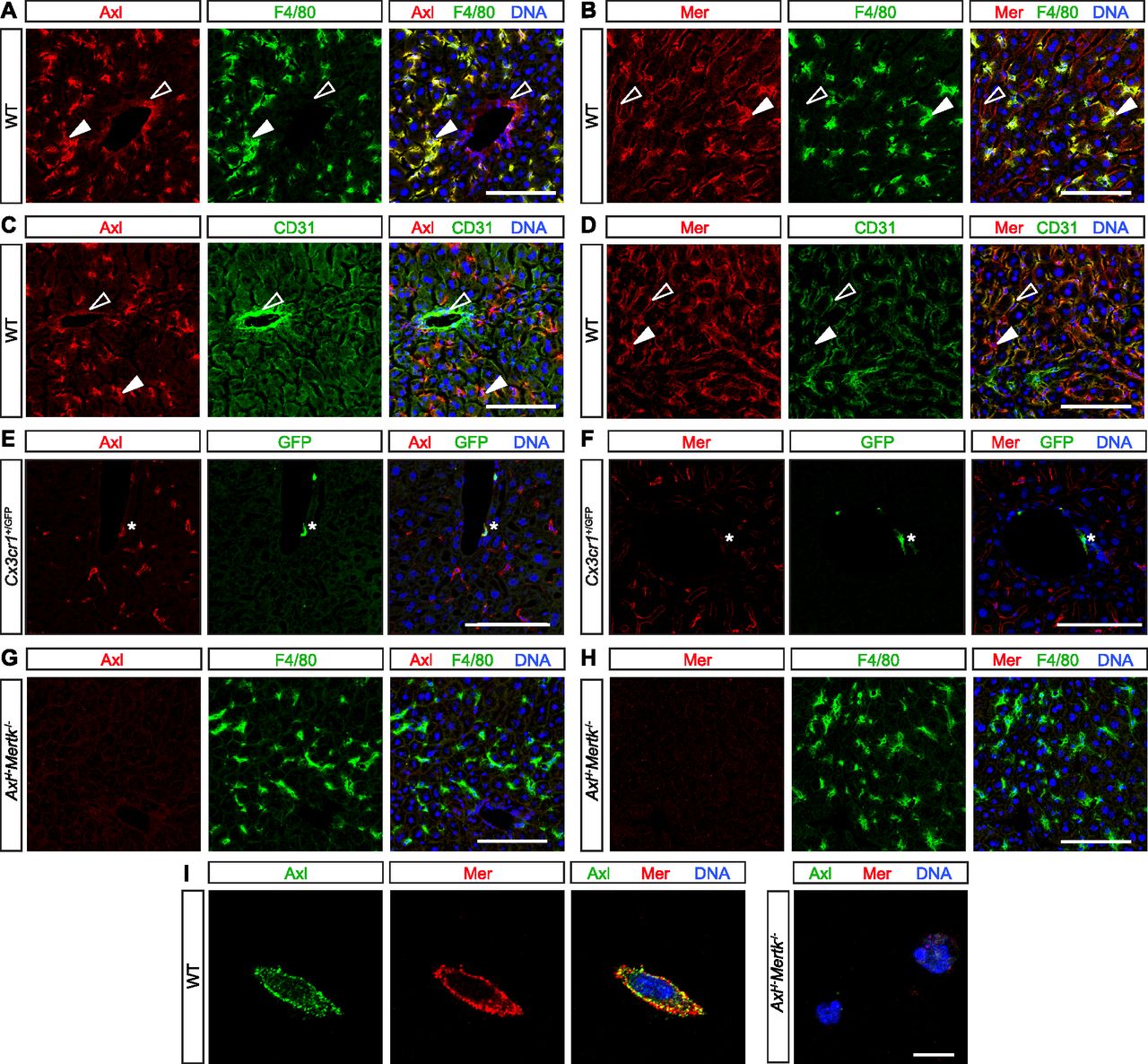
- Figure 1. TAM expression in normal liver.
(A, B, C, D, E, F, G, H) Mouse liver immunohistochemistry. (A, C, E) Axl (red) is expressed by F4/80+ Kupffer cells (KCs) (green, closed arrowheads) and strongly CD31+ blood-vessel-lining endothelial cells (green, open arrowheads) and by perivascular macrophages (E, green, asterisk). (B, D, F) Mer (red) is expressed by KCs (green, closed arrowheads), by weakly CD31+ sinus-lining endothelial cells (green, open arrowheads), and very weakly by perivascular macrophages (F, green, asterisk). (G, H) Mertk−/−Axl−/− mice were used for antibody controls. (I) Basal TAM expression on isolated KCs. Axl and Mer are co-expressed on the surface of individual cultured KCs. (See the Materials and Methods section for KC purification and culture). Bars (A, B, C, D, E, F, G, H): 100 μm, and (I): 10 μm.
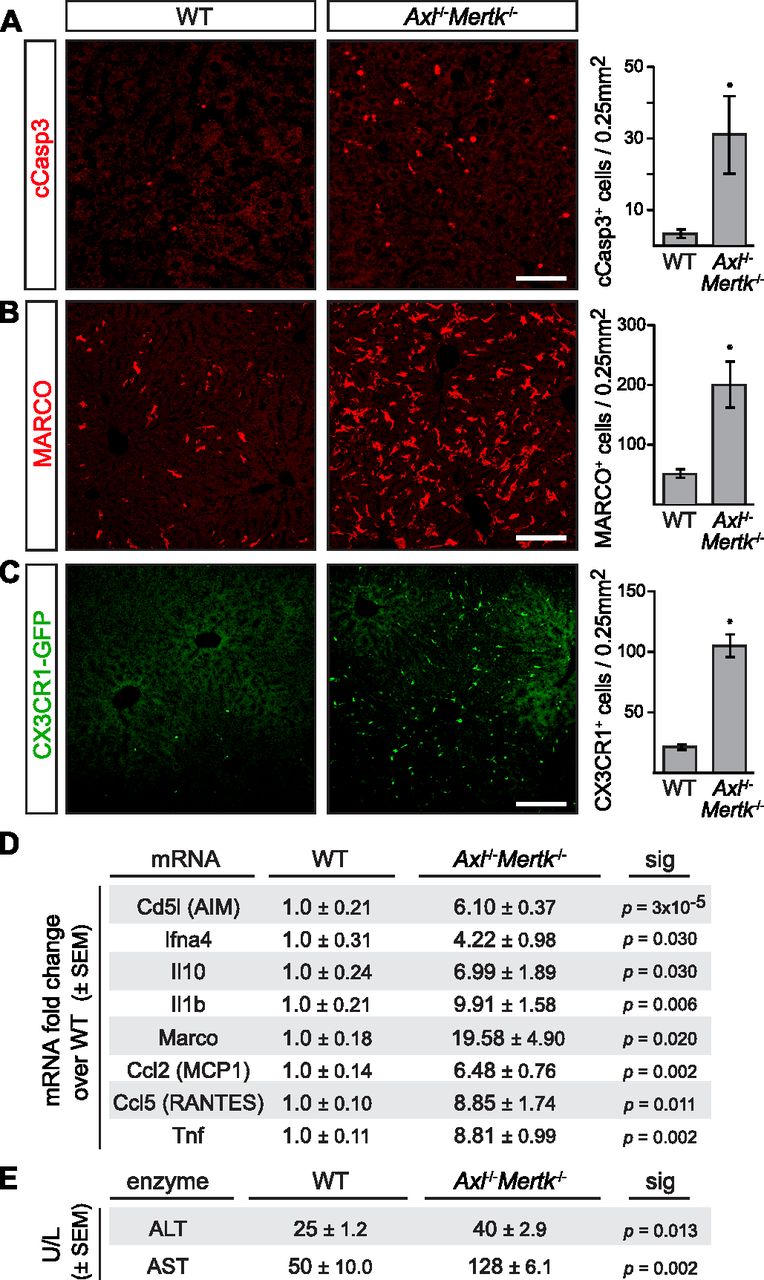
- Figure 2. Liver pathology in aged Axl−/−Mertk−/− mice.
(A) 9–12-mo-old Axl−/−Mertk−/− mice accumulate approximately ninefold more cCasp3+ apoptotic cells than WT mice. (B) 9–12-mo-old Axl−/−Mertk−/− exhibit an approximately fourfold increase in MARCO staining in liver than WT. (C) 9–12-mo-old Axl−/−Mertk−/−Cx3Cr1+/GFP mice have approximately fivefold more infiltrating GFP+ immune cells in liver than Cx3Cr1+/GFP. (D) Levels of the indicated mRNAs, isolated from 8 to 12 mo livers, and quantified by qRT-PCR relative to WT. (E) Serum ALT and AST activity from 7-mo-old WT and Axl−/−Mertk−/− mice. (A, B, C) Representative images from 3 to 4 mice per genotype. (D, E) Representative data from two independent experiments (n = 3–4 mice per genotype). Bars (A, B, C): 100 μm; *P < 0.05. t test.
- Figure S1. Consequences of aging in the liver.
(A) Representative H&E staining of WT (left) and Axl−/−Mertk−/− (right) liver at 12 mo. Arrowheads mark a pronounced increase in cellularity surrounding a blood vessel in the double mutants. Bars: 100 μm. (B, C, D, E, F) Indicated liver mRNAs measured by qPCR in mice of the indicated genotypes at 6–8 mo of age. Data points are results from separate mice; error bars are ± SEM. *P < 0.05, **P < 0.005, ****P < 0.0005. One-way ANOVA (Dunnett’s multiple comparison correction).
- Figure 3. Protective role of Mer in nonlethal Jo2 liver damage model.
(A) Extensive hemolysis (mottled darkening) of liver lobes 24 h after a nonlethal (0.3 mg/kg) Jo2 injection in exsanguinated (by cardiac puncture) Mertk−/− and Axl−/−Mertk−/−, but not in WT or Axl−/− mice. (B) Liver sections were analyzed by H&E staining (top row) and immunohistochemistry with indicated antibodies (bottom row). Markedly increased accumulation of apoptotic cells is observed in Mertk−/− and Axl−/−Mertk−/− liver. Bars: 100 μm. Arrowheads: damaged tissue and apoptotic cell accumulation. (A, B) Representative images from three experiments (n = 3–5 mice per genotype). (C) Apoptotic area quantification demonstrates a ∼14-fold increase in cCasp3+ cells. t test. (D, E) ALT/AST serum activity assays. n = 7–9 mice per genotype. *P < 0.05, **P < 0.01, ***P < 0.001. t test. (F) Liver lysates from WT mice injected with saline or Jo2 were immunoprecipitated for Mer and Axl and immunoblotted with indicated antibodies. (G) Quantification of the receptor phosphorylation (activation) results in (F). *P < 0.05, t test. Representative image from two experiments (n = 2 mice/treatment).
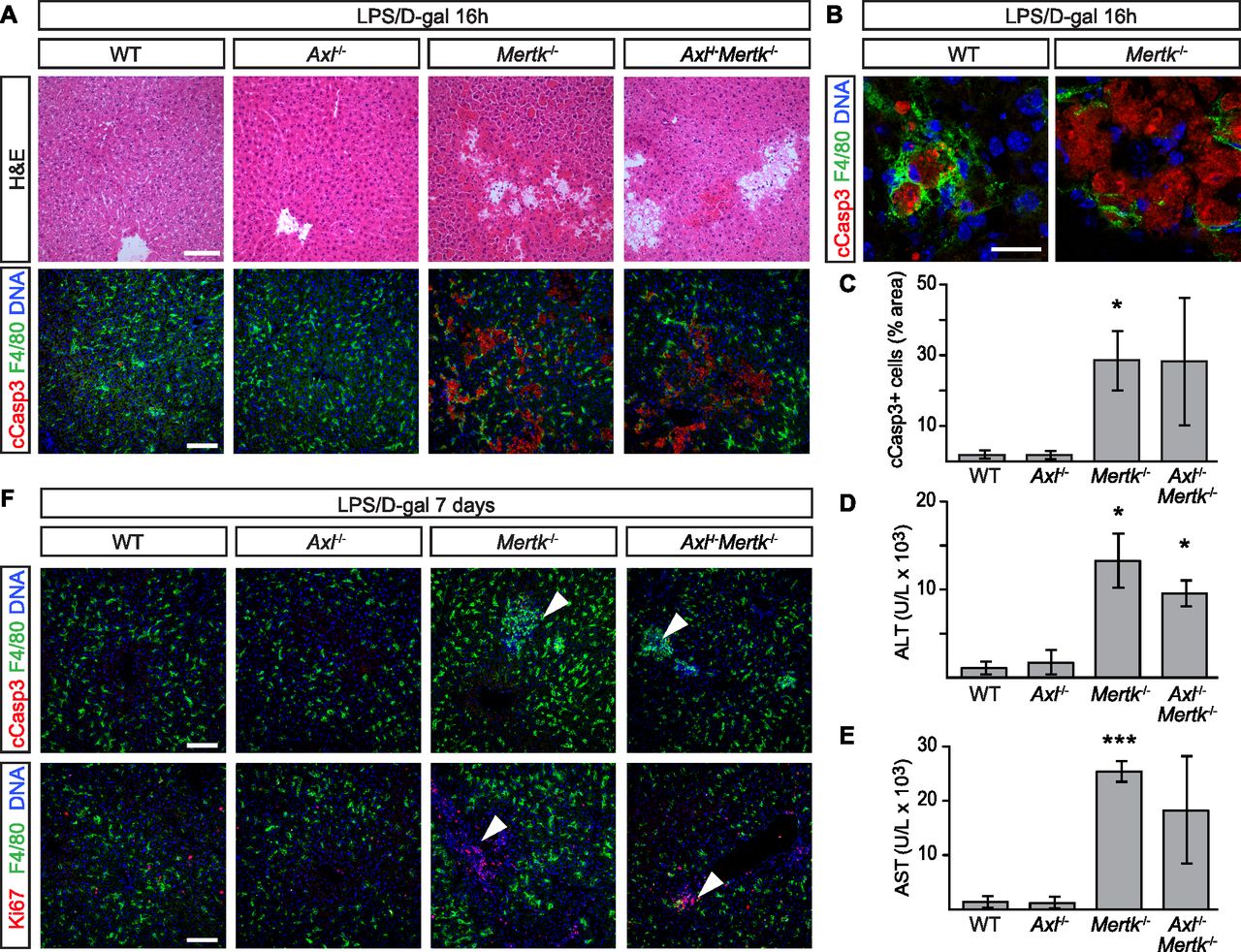
- Figure 4. Protective role of Mer in nonlethal LPS/D-gal liver damage model.
(A, B) Liver sections were analyzed 16 h after i.p. injection with 350 mg/kg D-gal and 1 μg/kg LPS by H&E staining and immunohistochemistry with the indicated antibodies. Increased accumulation of apoptotic cells can be observed in Mertk−/− and Axl−/−Mertk−/−. (A, B) Bars: 100 μm (A) and 20 μm (B). (B) shows that apoptotic cells are engulfed by F4/80+ Kupffer cells. (C) Apoptotic area quantification. (D, E) ALT and AST activity assays in sera 24 h postinjection. (F) Liver sections 7 d postinjection analyzed by H&E staining and immunohistochemistry with the indicated antibodies. Arrowheads in upper and lower rows represent unresolved loci of condensed, necrotic cCasp3+ cells, and proliferating Ki67+ cells, respectively, specifically in Mertk−/− and Axl−/−Mertk−/− livers. Bars: 100 μm. n = 3 per genotype, except Axl−/−Mertk−/− n = 2. *P ≤ 0.05, ***P ≤ 0.005. t test (C, D, E).
- Figure 5. Axl deletion from Kupffer cells (KCs).
(A) Liver sections from tamoxifen-treated (+Tx) Cx3cr1CreER/+Axlf/fMertkf/f mice and constitutive Cx3cr1Cre/+Axlf/fMertkf/f mice stained with antibodies to F4/80 (top row, green) and Axl (middle row, red), illustrating Cx3cr1Cre/+-driven loss of Axl from KCs but not endothelial cells (ECs) in constitutive Cx3cr1Cre/+Axlf/fMertkf/f mice, but normal strong Axl expression in KCs in Tx-treated Cx3cr1CreER/+Axlf/fMertkf/f mice. Bottom row is the merged image of top and middle rows, with DNA visualized by Hoechst 33258 (blue). Note that Axl immunostaining in KCs of the constitutive Cx3cr1Cre/+Axlf/fMertkf/f liver is dramatically reduced but not completely eliminated. Examples of KCs with low residual Axl staining are indicated by asterisks. Axl immunostaining in ECs (arrowheads) persists in both genotypes. (A, B) Same analyses as in (A), except that middle row sections are stained with a Mer antibody. Mer immunostaining in ECs (arrowheads) persists in both genotypes. (C) Peritoneal macrophages from Cx3cr1Cre/+Axlf/fMertkf/f mice, ±treatment with 10 nM Gas6, were immunoprecipitated and blotted for Axl. The low residual level of Axl immunostaining in KCs of these mice (A) may be due to the presence of a low level of a truncated Axl (asterisk) produced by Cre-mediated recombination. (D) Levels of the indicated mRNAs, isolated from 6 to 7 mo livers of the indicated genotypes, and quantified by qRT-PCR. t test. (E) Cleaved Casp3+ apoptotic cell accumulation in the liver 24 h after a nonlethal Jo2 injection in Tx-injected Cx3cr1CreER/+Axlf/fMertkf/f mice, in which Axl and Mer expression in KCs is maintained (left), versus Cx3cr1Cre/+Axlf/fMertkf/f mice, in which Axl and Mer expression in KCs is lost (right). Bars (A, B, E): 100 μm.
- Figure 6. Protective role of Axl in APAP intoxication.
(A) Representative images showing extensive hemorrhage and congestion of liver lobes 48 h after administration of acetaminophen (APAP; 300 mg/kg) to Axl−/− but not WT or Mertk−/− mice. These in situ liver images are from non-perfused mice. (B) Representative liver sections from 48 h APAP-treated mice were analyzed by H&E staining. Markedly increased congestion and blood hemorrhage is observed in Axl−/− but not WT or Mertk−/− liver. Bar: 100 μm. (C) Measurement of circulating ALT (U/l, units per liter) 48 h after APAP administration in mice of the indicated genotypes. (D) Immunostaining for cleaved Casp3+ cells in liver sections of WT and Axl−/− mice at 48 h after APAP administration. Bar: 50 μm. (E) Induction of soluble Axl (sAxl) in WT and Mertk−/− mice 48 h after APAP. The trend toward sAxl induction in Mertk−/− mice is not statistically significant. As for all APAP treatments, mice were fasted for 16 h before drug administration. (F) Levels of the indicated mRNAs, isolated livers of the indicated genotypes 48 h after APAP treatment, and quantified by qRT-PCR. *P < 0.05; **P < 0.005. Two-way ANOVA (Bonferroni multiple comparison correction) (C, E); t test (F).
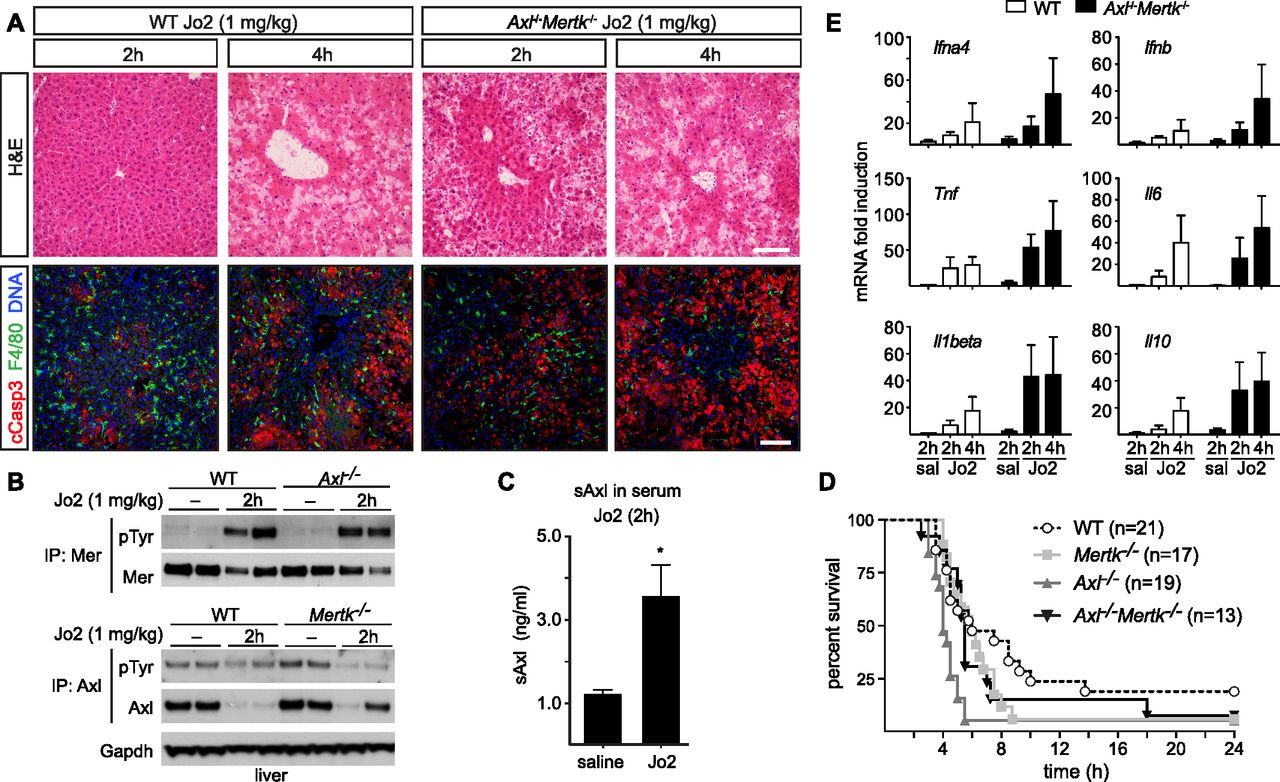
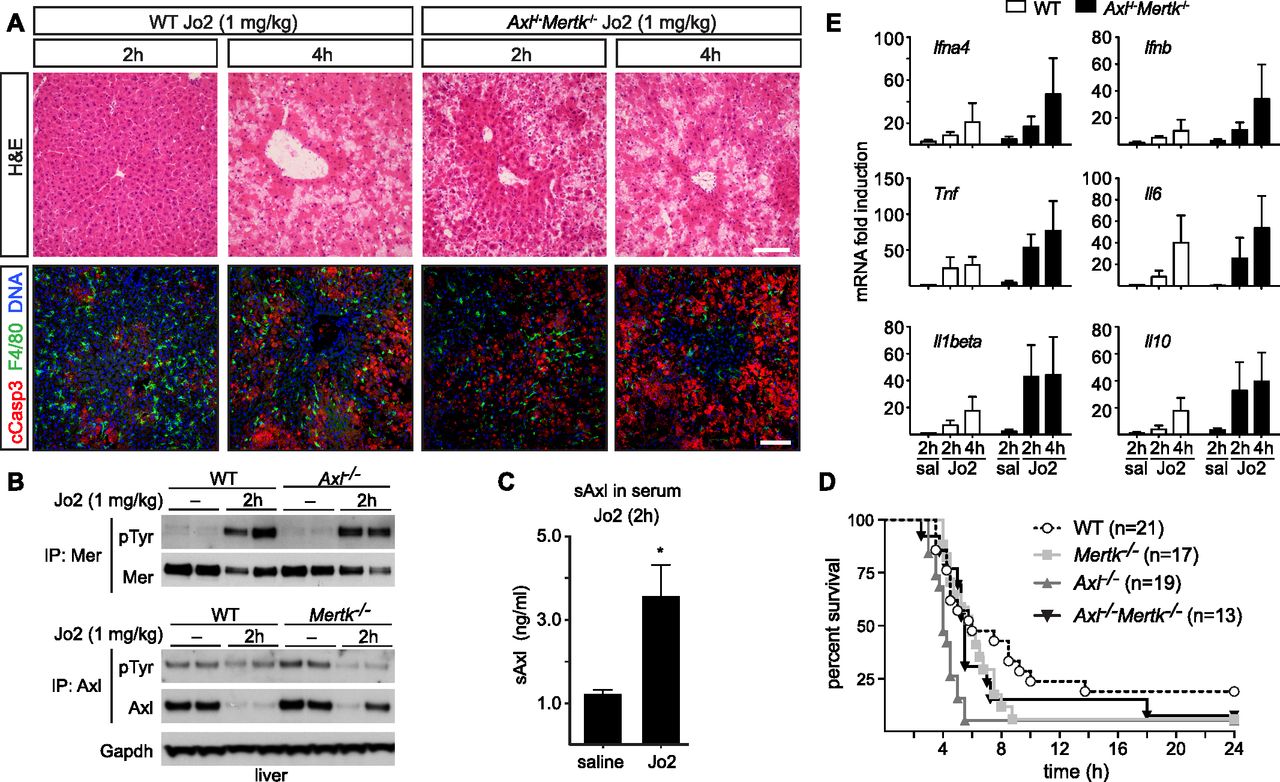
- Figure 7. Protective role of Axl and Mer in lethal Jo2 liver damage model.
(A) Liver sections at 2 and 4 h after a lethal i.p. injection (1 mg/kg) of Jo2 were analyzed by H&E staining and immunohistochemistry with indicated antibodies. Increased accumulation of apoptotic cells can be observed in Axl−/−Mertk−/−. Bars: 100 μm. Representative images from three experiments (n = 2–5 mice per genotype). (B) Liver lysates from WT and Axl−/− mice injected with saline or Jo2 (1 mg/kg) were immunoprecipitated for Mer and Axl and immunoblotted with indicated antibodies; n = 2, each lane an individual mouse. (C) sAxl ELISA in serum of saline and Jo2 (1 mg/kg) injected mice; n = 2, *P < 0.05. t test. (D) Survival of mice of the indicated genotypes after Jo2 (1 mg/kg) injection. (E) Expression of the indicated inflammatory markers was analyzed by qRT-PCR from liver mRNA samples; representative results from one to three experiments.
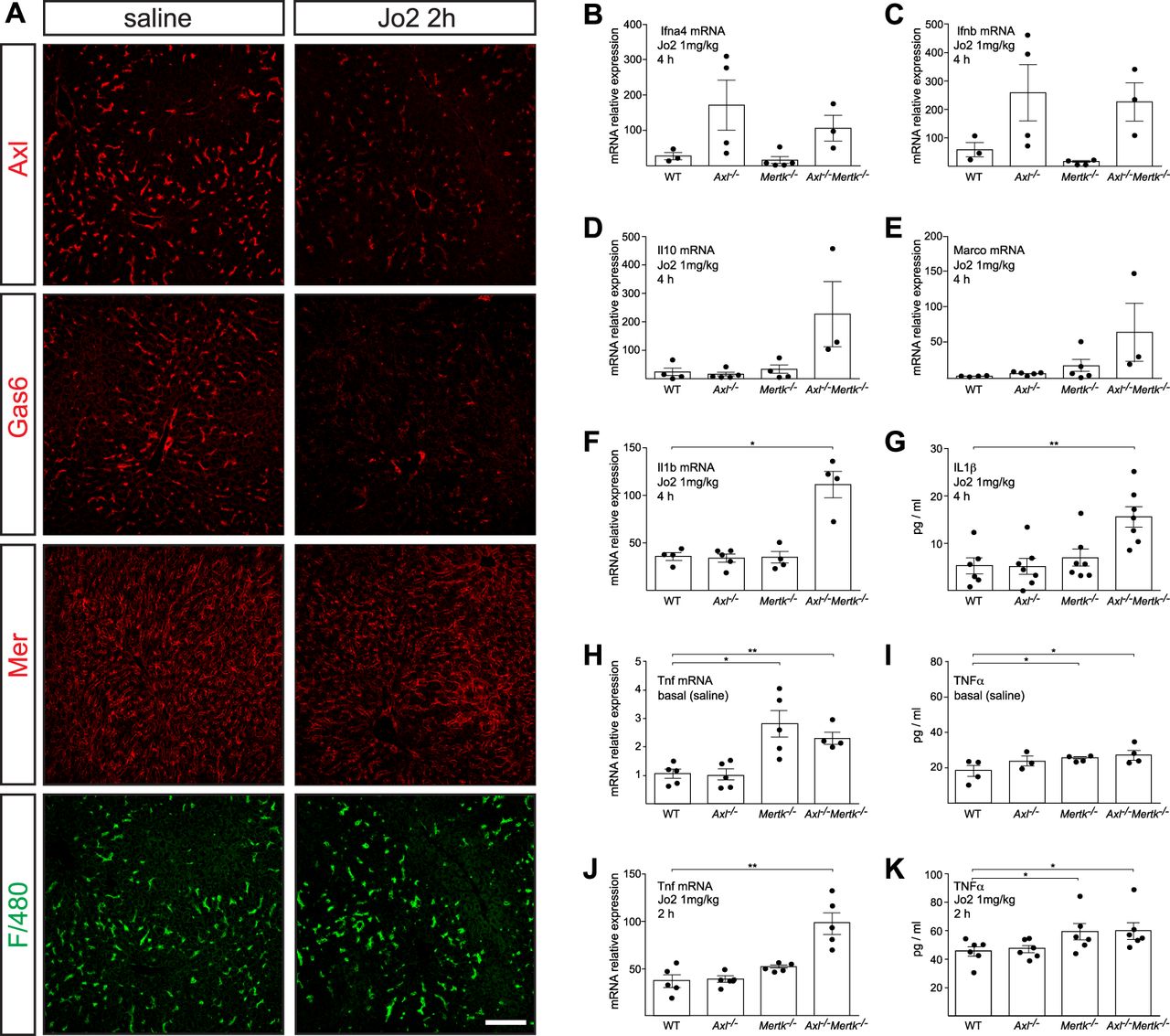
- Figure S2. Hepatic responses to high (lethal) dose Jo2.
(A) Representative examples of expression of the indicated proteins in the liver, as detected by immunohistochemistry, at 2 h after saline (left panels) or lethal Jo2 (right panels). Bars: 100 mm. (B, C, D, E, F, G, H, I, J, K). The indicated mRNAs (B, C, D, E, F, H, J) and proteins (G, I, K) were measured by qPCR and ELISA, at 2 or 4 h (as indicated) after an IP injection (1 mg/kg) of Jo2 into mice of the indicated genotypes. mRNAs for qPCR were isolated from the liver; proteins for ELISA were from serum. Data points are results from separate mice; error bars are ± SEM. *P ≤ 0.05, **P ≤ 0.005. t test (B, C, D, E, F, G, H, I, J, K).
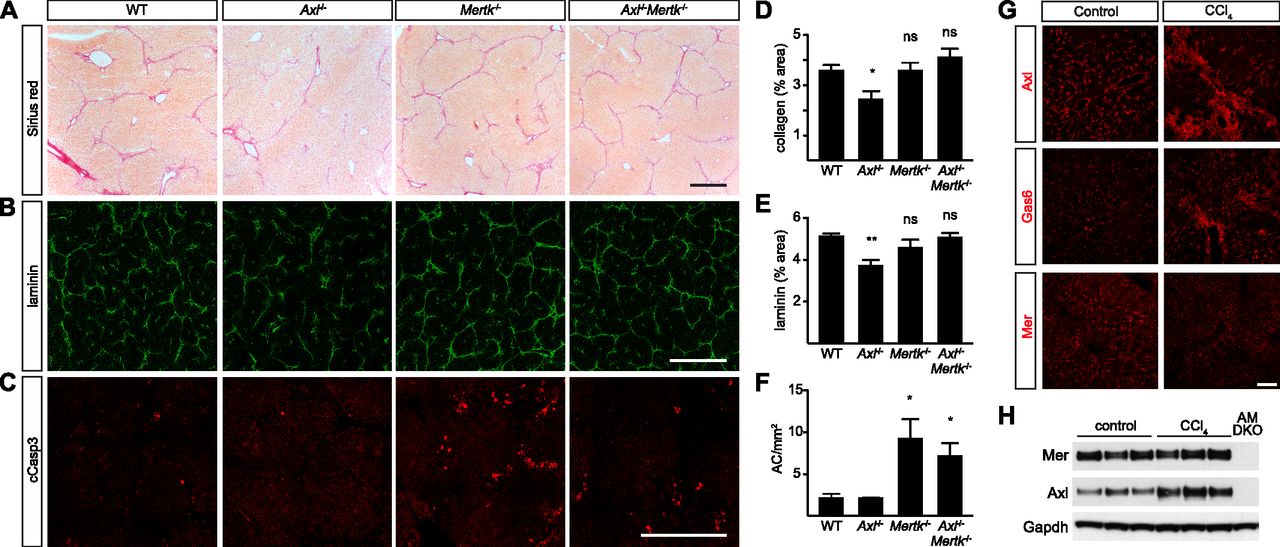
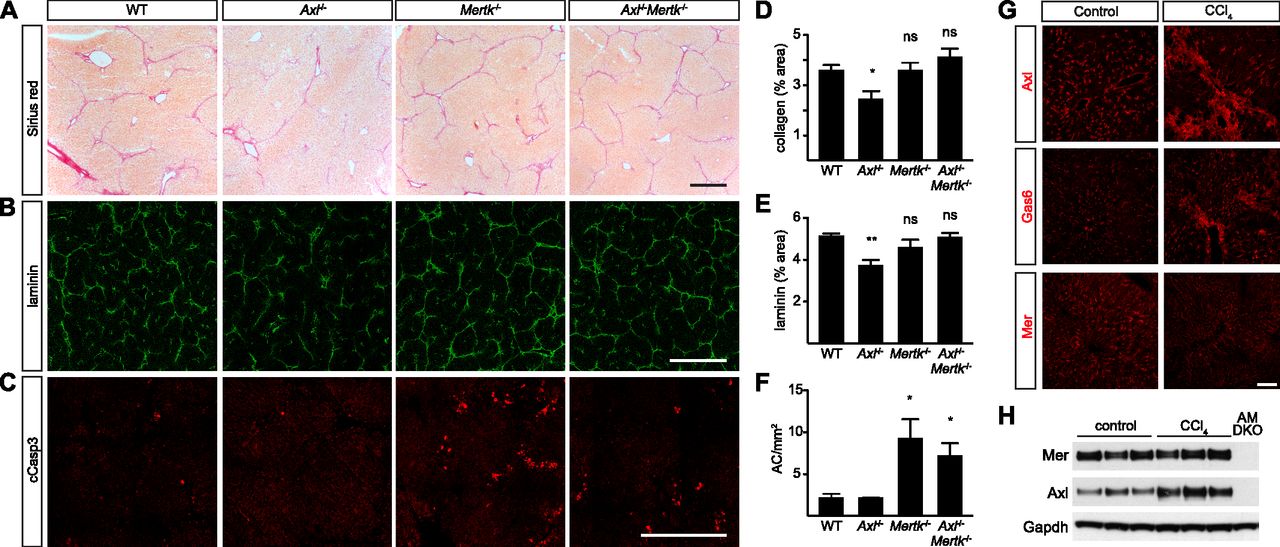
- Figure 8. Axl promotion of CCl4-induced liver fibrosis.
(A) Sirius red staining, showing collagen deposition, of liver sections from mice of the indicated genotypes, injected with CCl4 three times per week for 6 wk. (B) Laminin staining of similar sections. (C) Cleaved caspase 3 (cCasp3) staining of similar sections. (A, B, C, D, E, F) Quantification of the results in (A, B, C), respectively. (G) Up-regulation of Axl and Gas6, but not Mer, in fibrotic liver. (H) Western blot showing increased expression of Axl in fibrotic liver. (A, B, C, G) Bars (A, B, C): 0.5 mm (G) and 100 μm. Representative images from six mice per genotype. *P < 0.05, **P < 0.01. t test (D, E, F).
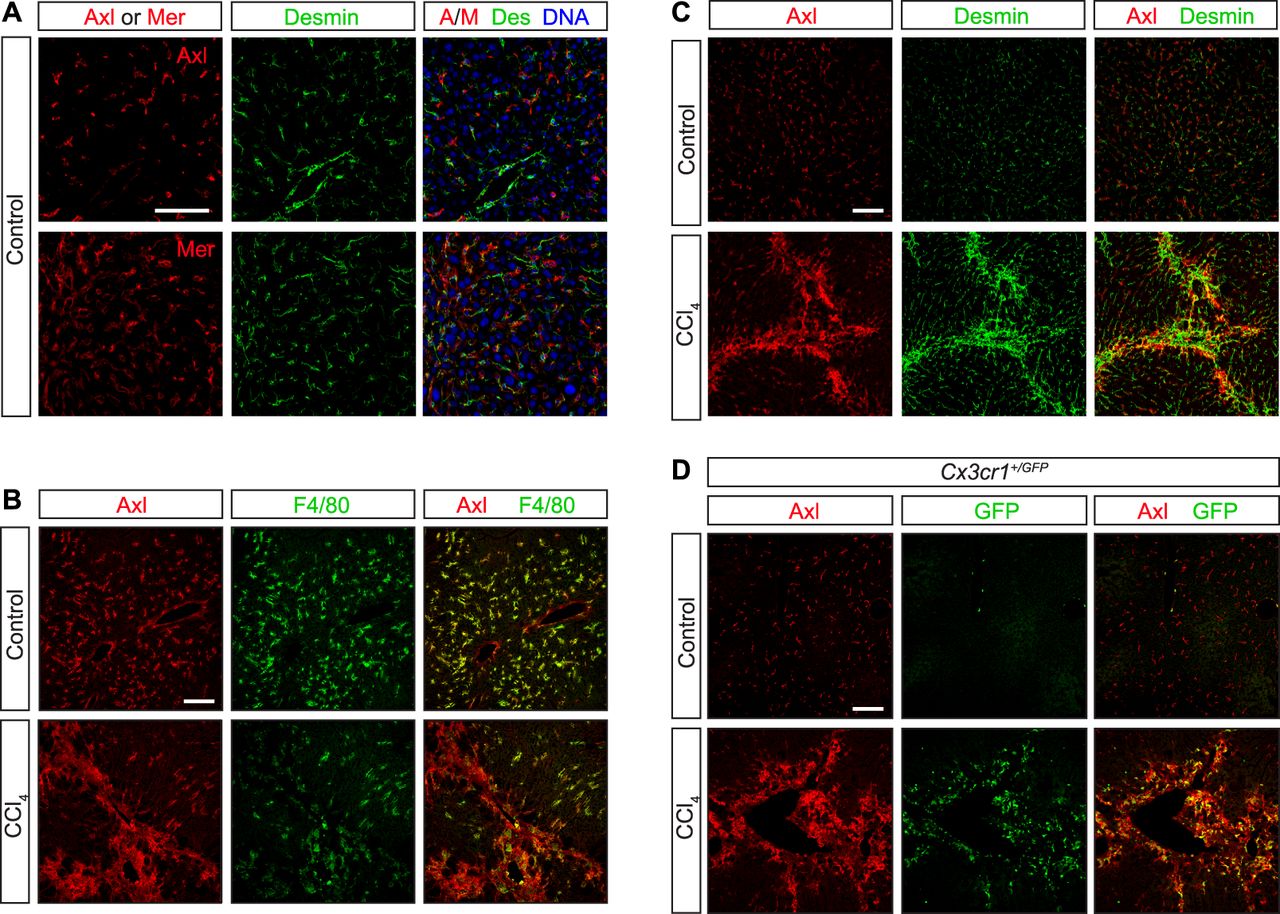
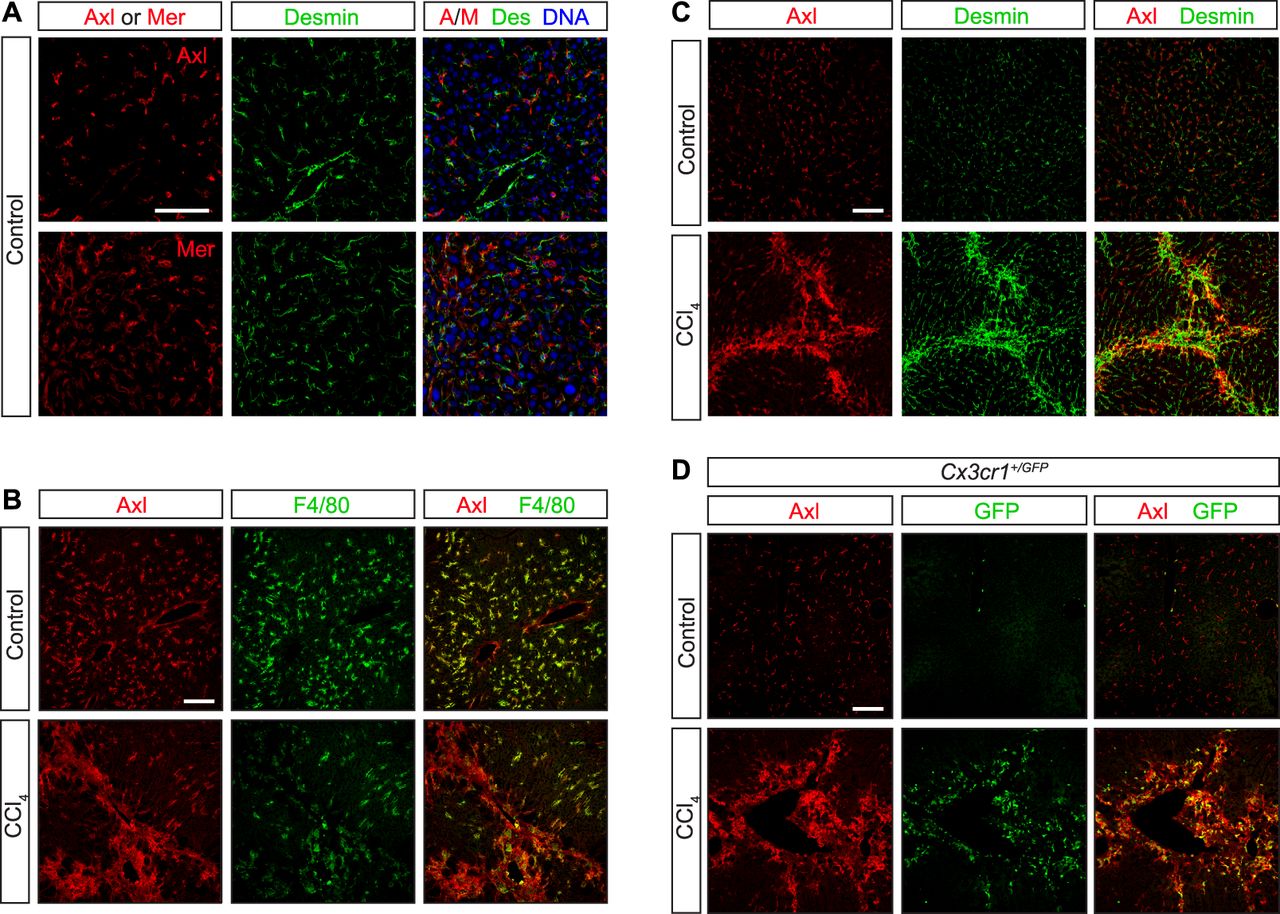
- Figure S3. Axl expression in CCl4 liver fibrosis model.
WT and Cx3cr1+/GFP mice were injected with CCl4 three times per week for 6 wk. (A, C) Neither Axl nor Mer co-localizes with the hepatic stellate cell marker desmin under basal conditions—before CCl4 treatment (A, C, control). (B) As is also shown in Fig S1A and B; these receptors are instead prominently expressed in F4/80+ Kupffer cells (B). (C, D) After CCl4 treatment, however, partial overlap between desmin+ and Axl+ cells is observed (C), although a stronger overlap is seen between Axl and infiltrating CX3CR1+ cells (D). Bars: 100 μm.





Supplementary Materials




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- TAM receptors control actomyosin dynamics in osteoclasts via RHOA-COFILIN-MYOSIN II signaling
- Distinct circulating monocytes up-regulate CD52 and sustain innate immune function in patients with cirrhosis unless acute decompensation emerges
- Tissue-specific modifier alleles determine Mertk loss-of-function traits
- Clonal expansion and activation of tissue-resident memory-like TH17 cells expressing GM-CSF in the lungs of patients with severe COVID-19