




Article Figures & Data
Figures
- Figure 1. Chk1 inhibitors reduce expression of Chk1 and induce DNA damage.
(A) λ820 mouse lymphoma cells were treated with LY2603618 for 24 h, and cells were analyzed by immunoblotting for Chk1 and loading control β-actin. (B) λ820 mouse lymphoma cells were treated with CCT245737 and cells were analyzed by immunoblotting for apoptosis marker cleaved PARP, total or phosphorylated (p)Chk1 and DNA damage marker phosphorylated histone 2AX (γH2Ax). (C) λ820 mouse lymphoma cells were treated with LY2603618 or ATR inhibitor VE821 for 24 h, and cells were analyzed by immunoblotting for total Chk1, pChk1, phosphorylated (p)RPA2, and loading control β-actin.
Source data are available for this figure.
Source Data for Figure 1[LSA-2020-00671_SdataF1.zip]
- Figure 2. Generation of a kinase-dead Chk1 mouse model.
(A) Clustering analysis of Chk1 from yeast, human, mouse, and frog. Arrow indicated aspartate 130 (D130). (B) Targeting strategy to generate a kinase-dead Chek1 allele. Top part of the mouse Chek1 gene. Middle part of the Chek1 gene showing targeted allele in ES cells. Bottom part of the Chek1 gene showing the Chek1 gene after removal of neo cassette in an Flp-deleted mouse. (C) PCR genotyping of three offspring after mating between a heterozygous Chek1 knockin mouse (Chek1D130A/wt). (B) PCR primers (NDEL1 and 2) are shown in (B). (D) Results of genotyping of embryos or offspring from time pregnancies between Chek1D130A/wt females and Chek1D130A/wt males. (E) Results of genotyping of offspring from crosses of heterozygous Chek1 knockin males or females to C57BL/6 mice.
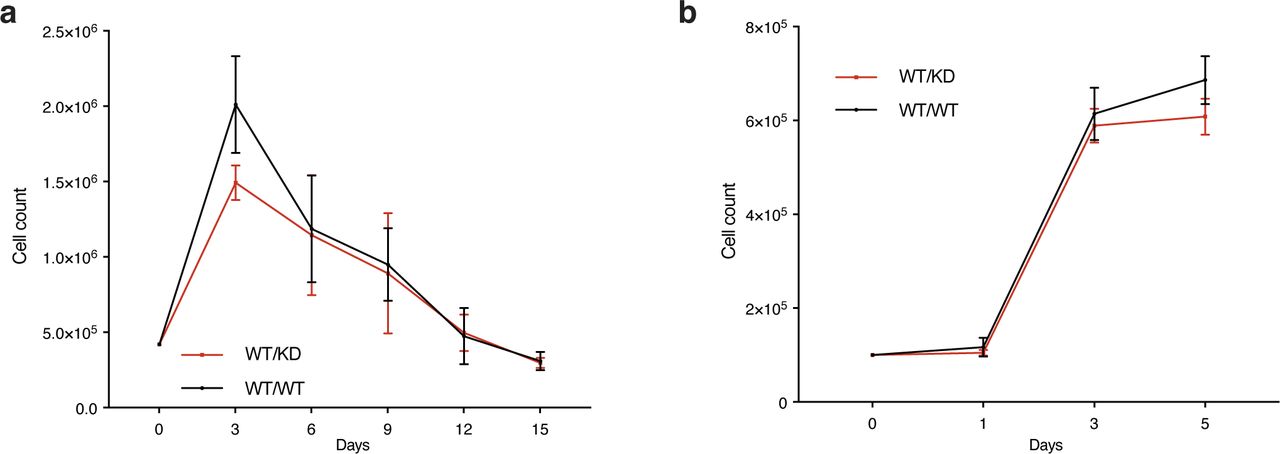
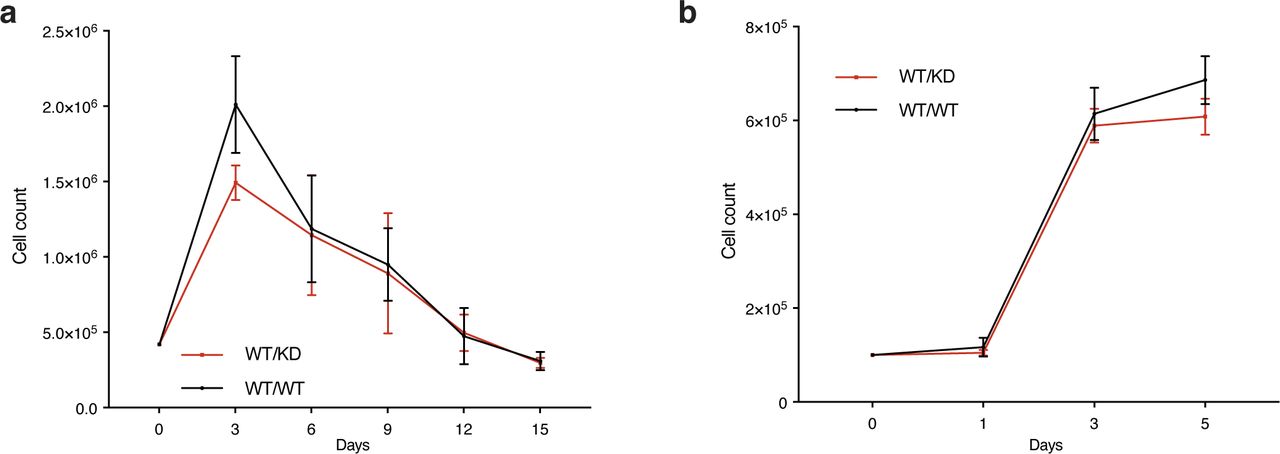
- Figure S1. Heterozygosity of a kinase-dead Chk1 expressing allele does not impact growth of MEFs.
(A) Timed pregnancies between Chek1D130A/+ mice and C57BL/6 mice were used to make MEFs. Passage 3 MEFs were seeded at 0.5 million cells in a 5-cm plate and cultured for 3 d. Cells were counted, and 0.5 million cells were plated and cultured for 3 d again. The procedure was continued until the cells no longer were able to proliferate in 3 d. (B) Growth curves of MEFs grown for 5 d.
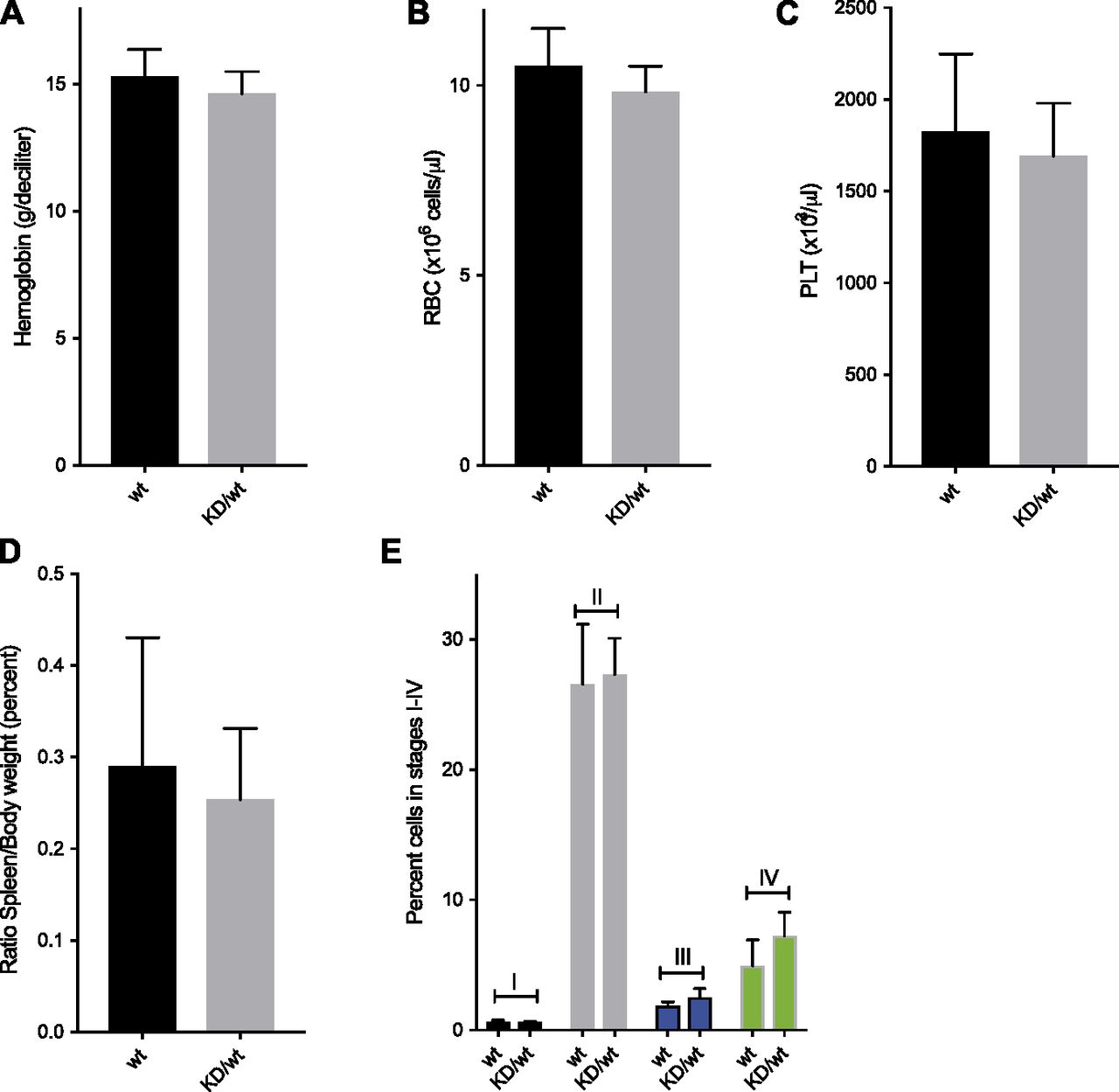
- Figure 3. Heterozygous Chek1 knockin mouse mice do not have a problem with erythropoiesis.
(A, B, C, D) 21 wild-type (wt) and 15 heterozygous Chek1 knockin (KD/wt) 45–60-wk-old mice were analyzed for hemoglobin content (A), red blood cell count (B), platelet count, and spleen weight (D). (E) Six wt and three KD/wt mice were analyzed for different stages (I–IV) of erythroid development using Ter119/CD71 staining and flow cytometry. Representative flow cytometry plots are seen in Fig S2.
- Figure S2. Bone marrows from wild-type and Chek1KD/w mice were analyzed for different stages (I–IV) of erythroid development using Ter119/CD71 staining and flow cytometry.
Shown are representative flow cytometry plots and the data from multiple mice is seen in Fig 3E.
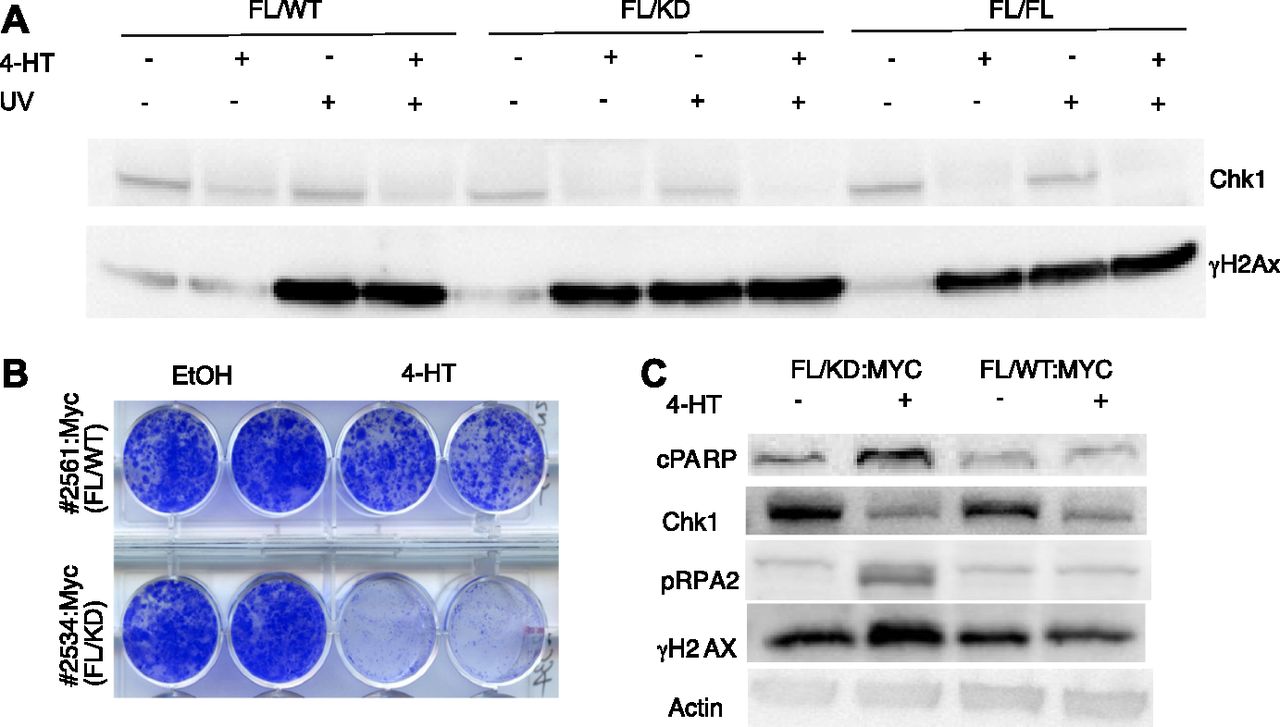
- Figure 4. Chk1 kinase activity is essential to suppress DNA damage signaling activation and for fibroblast viability.
(A) Mouse fibroblasts from CreER mice carrying one floxed Chek1 allele and a wild-type (FL/WT), kinase-dead (FL/KD), or another floxed allele (FL/FL) were treated with vehicle or 4-hydroxytamoxifen (4HT) for 4 d and/or with UVB for 30 s (∼700 J/m2) and harvested 2 h after radiation or mock. Immunoblotting was performed for Chk1 and γH2Ax. (B) Mouse fibroblast expressing CreER, and a floxed and a kinase-dead or wild-type Chek1 allele were transduced with a Myc-expressing retrovirus. Cells were treated with vehicle or tamoxifen and long-term survival was determined by staining cell monolayers. (A, C) Cells manipulated the same as in (A) were analyzed by immunoblotting for indicated markers of apoptosis (PARP), replication stress (threonine 21 phosphorylated-RPA2), and DNA damage (γH2Ax).
- Figure 5. Expression of only a kinase-dead allele of Chek1 suppresses lymphoma growth.
(A) Kaplan–Meier survival analysis of λ-Myc; Cdkn2a+/− mice carrying the indicated Chek1 genotypes. (A, B, C, D) Lymphomas from mice generated from mice in (A) were transplanted into eight C57BL/6 mice per indicated genotype. When white blood counts reached between 20 and 50 × 103 cells/μl, four mice received vehicle or tamoxifen by i.p. injection. White blood count was followed until relapse or when palpable tumors required euthanasia of the animals.
- Figure S3. λ-Myc lymphomas of indicated Chek1 genotypes were transplanted into C57BL/6 recipient mice.
When white blood count was above normal, mice were treated with tamoxifen to induce Cre-mediated deletion of the floxed Chek1 allele. Tumors were harvested from the mice when becoming palpable and protein and DNA were extracted. (A) Western blot analysis of protein from indicated lymphomas. The blots were probed with antibodies against human Myc to confirm that the lymphoma were λ-Myc lymphomas, with anti-Chk1 to investigate if Chk1 had been deleted, with anti-γH2ax to analyze if DNA damage was induced and with anti–β-actin as a loading control. (B) PCR analysis with primers spanning one of the loxP sites in the genome of floxed mice. The larger band disappears if floxing occurs. (C) PCR analysis with primers detecting a DNA sequence that is created if Cre recombines in the genome of mice with floxed Chek1 alleles.
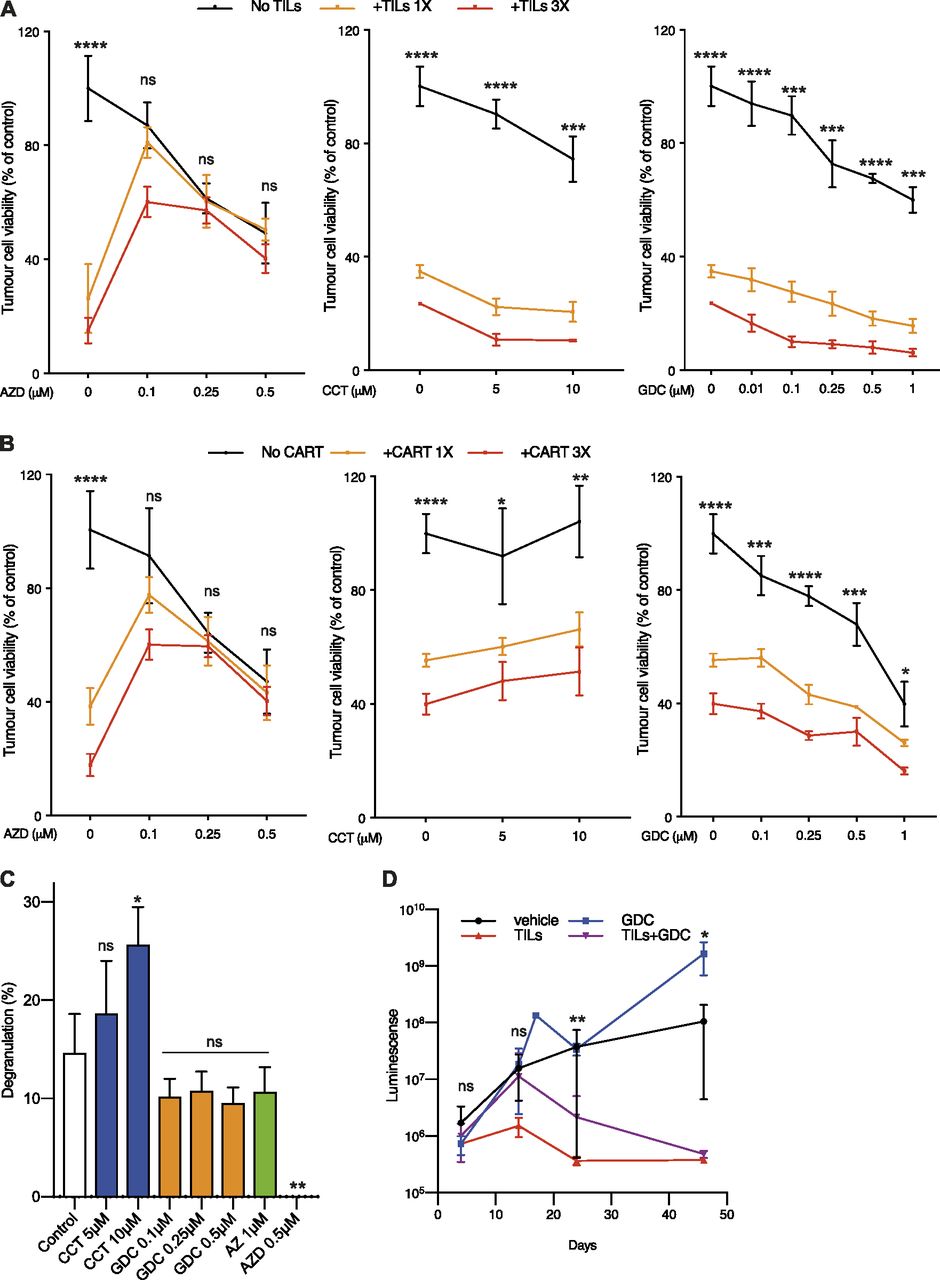
- Figure 6. Chk1 inhibition does not interfere with T-cell–mediated killing of melanoma cells.
(A) 33F8-luciferase expressing melanoma cells were treated with indicated Chk1 inhibitors in the absence or presence of autologous TILs (1:1 or 3:1 TILs vs melanoma cells). Luciferin was added and viability was measured in a 96-well plate luminometer. (A, B) The same experimental setup as in (A) but using HER2-directed CAR-T cells instead of TILs. (C) 33F8 cells were mixed with autologous TILs in the presence of indicated inhibitors. Surface expression of CD107a as a marker of T-cell degranulation was determined by flow cytometry. (D) NOD/SCID/IL2 receptor gamma knockout mice transgenic for human interleukin-2 (hIL2-NOG mice) were transplanted with 33F8-luciferase melanoma cells. When tumors were detectable by IVIS imaging, TILs were injected and mice were dosed in indicated groups of animals (n = 4). Mice were followed with IVIS imaging until tumor size reached the ethics limit (10 mm on the shortest side). Shown are median IVIS signal and standard error of the mean.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Related Articles
Cited By...
- No citing articles found.