



Abstract
Angiogenesis and lymphangiogenesis are key processes during embryogenesis as well as under physiological and pathological conditions. Vascular endothelial growth factor C (VEGFC), the ligand for both VEGFR2 and VEGFR3, is a central lymphangiogenic regulator that also drives angiogenesis. Here, we report that members of the highly conserved BACH (BTB and CNC homology) family of transcription factors regulate VEGFC expression, through direct binding to its promoter. Accordingly, down-regulation of bach2a hinders blood vessel formation and impairs lymphatic sprouting in a Vegfc-dependent manner during zebrafish embryonic development. In contrast, BACH1 overexpression enhances intratumoral blood vessel density and peritumoral lymphatic vessel diameter in ovarian and lung mouse tumor models. The effects on the vascular compartment correlate spatially and temporally with BACH1 transcriptional regulation of VEGFC expression. Altogether, our results uncover a novel role for the BACH/VEGFC signaling axis in lymphatic formation during embryogenesis and cancer, providing a novel potential target for therapeutic interventions.
Introduction
The blood and lymphatic networks are two evolutionarily conserved transport systems that provide complementary functions in the maintenance of tissue homeostasis. In particular, the formation of new blood and lymphatic vessels is a prerequisite for vertebrate embryonic and postnatal development. During embryogenesis, the blood circulatory system is first to emerge. As early as mouse embryonic day (E) 7.5, vasculogenesis, the formation of a primitive vascular plexus through proliferation, migration, and differentiation of endothelial cells (ECs), takes place. Later on, this plexus undergoes massive remodeling via angiogenesis, which involves also arteriovenous differentiation (Chung & Ferrara, 2011). At E10.5, a distinct subpopulation of ECs within the cardinal vein commits to the lymphatic lineage, buds off, and migrates to establish primitive lymph sacs, which through further sprouting, give rise to the peripheral lymphatic system (Oliver & Alitalo, 2005; Semo et al, 2016). During adult life, most blood and lymphatic vessels are quiescent, with the exception of female reproductive organs during the ovarian cycle and the placenta during pregnancy. Reactivation of angiogenesis and lymphangiogenesis, however, is a hallmark of pathological processes associated with wound healing, myocardial infarction, allograft rejection, chronic inflammation, tumor progression, and malignant cell dissemination (Oliver & Alitalo, 2005; Chung & Ferrara, 2011).
The VEGF family of growth factors and its receptors are central signaling pathways, controlling angiogenesis and lymphangiogenesis during development and adulthood. VEGFs exert their activity by binding to the tyrosine kinase receptors (vascular endothelial growth factor receptor-1) VEGFR-1, VEGFR-2, and VEGFR-3 expressed in blood and lymphatic endothelial cells (LECs) (Lohela et al, 2009). VEGFA, the ligand for VEGFR-1 and VEGFR-2, is best known for its role in angiogenesis, stimulating EC proliferation and migration and increasing vascular permeability. VEGFB, which also binds to VEGFR1, plays a role in cell survival and indirectly promotes VEGFA-induced angiogenesis (Lal et al, 2018).
Vascular endothelial growth factor C (VEGFC) is another critical player in VEGF signaling. This protein, which signals through the receptors VEGFR-3 and VEGFR-2, plays a key role across species, guiding both lymphatic and blood vasculature development and remodeling (Karkkainen et al, 2004; Kuchler et al, 2006; Yaniv et al, 2006; Lohela et al, 2009; Hogan et al, 2009a; Gore et al, 2011; Villefranc et al, 2013; Shin et al, 2016). VEGFC is expressed by endothelial and non-endothelial cells, thus acting via both autocrine and paracrine signaling (Covassin et al, 2006b; Kodama et al, 2008; Lohela et al, 2008; Khromova et al, 2012; Helker et al, 2013; Villefranc et al, 2013). Evidence for its necessity for proper lymph formation is revealed by the finding that ectopic expression of Vegfc in mouse ear or skin keratinocytes results in hyperplasia of lymphatic vessels. Similarly, mice and zebrafish lacking VEGFC fail to develop a lymphatic vasculature as initial sprouting and directed migration of lymphatic progenitors from the cardinal vein are arrested (Karkkainen et al, 2004; Kuchler et al, 2006; Yaniv et al, 2006; Villefranc et al, 2013; Shin et al, 2016). In addition to its predominant role in lymphangiogenesis, VEGFC was shown to induce angiogenesis upon overexpression in the mouse cornea, skin, or ischemic hind limb. Moreover, coronary vessel development is VEGFC dependent, and hearts devoid of VEGFC reveal dramatic delay in the formation of subepicardial sprouts (Chen et al, 2014). Recently, it has been shown by in vitro studies that the let-7a/TGFBR3 axis regulates angiogenesis through transcriptional regulation of VEGFC (Wang et al, 2019).
VEGFC has a critical role not only during development but also during tumor progression. In various human cancers, enhanced expression of VEGFC and higher levels of VEGFC in serum are commonly associated with tumor aggressiveness and lymph-node metastasis (Su et al, 2007; Lohela et al, 2009). In esophageal carcinoma, for instance, angiogenesis is driven via the phosphoinositide-phospholipase C-ε (PI-PLCε)/NF-κB signaling pathway by direct promotion of VEGFC transcription (Chen et al, 2019). In xenograft or transgenic tumor models, stimulation of lymphangiogenesis by VEGFC promotes malignant cell dissemination (Stacker et al, 2014). Furthermore, blockade of Vegfc expression in tumor cells by stably transfected small interfering RNAs reduces lymphangiogenesis and lymph node metastasis of murine mammary cancers. Similarly, soluble VEGFR-3 protein has been shown to inhibit VEGFC-induced tumor lymphangiogenesis and metastatic spread in a breast cancer mouse model (Wissmann & Detmar, 2006).
A series of environmental and cellular factors were shown to modulate VEGFC expression. For instance, IL-6 and IL-17 regulate VEGFC expression via the PI3K-Akt or extracellular-signal-regulated kinase (ERK) 1/2 pathways, whereas MicroRNA-1826 significantly down-regulates VEGFC expression in human bladder cancer (Chen et al, 2012). Vegfc mRNA levels are heightened in the adipose tissue of obese mice, pointing to adipocytes as a source of elevated VEGFC levels in obesity (Karaman et al, 2015). In adults, inflammation induces robust up-regulation of VEGFC expression by macrophages (Baluk et al, 2005). In addition, various hormones, transcription factors, metallothioneins, and microenvironmental stresses (e.g., hyperthermia, oxidative stress, and high salt) were shown to control VEGFC expression (Cohen et al, 2009; Machnik et al, 2009; Sapoznik et al, 2009; Chen et al, 2012; Schuermann et al, 2015; Gauvrit et al, 2018).
Here, we report that BACH transcription factors, which are known to be involved in various intracellular signaling pathways, modulate VEGFC expression during embryonic development and tumor progression, offering a better understanding of blood and lymph vessel formation during physiological and pathological conditions.
Results
Spatial and temporal expression of bach2 during zebrafish development
To explore the molecular mechanisms governing VEGFC expression, we searched for genes that co-express with VEGFC in human prostate cancer datasets (Glinsky et al, 2004). We identified 434 genes, on which we further applied the Promoters of Clusters analysis (Tabach et al, 2007). This method screens the promoters of genes with shared biological function against a library of transcription factor–binding motifs and identifies those which are statistically overrepresented. Five factors passed the basic significance threshold, out of which only two possessed binding sites within the VEGFC promoter. We then searched for evolutionarily conserved regions, which often represent potential DNA regulatory elements (Ovcharenko et al, 2004). Comparison of the VEGFC promoters of human, mouse, and zebrafish revealed two highly conserved BACH (broadcomplex-tramtrack-bric-a-brac [BTB] and cap’n’col-lar type of basic leucine zipper [CNC-bZip] homology)-binding sequences (Fig 1A).
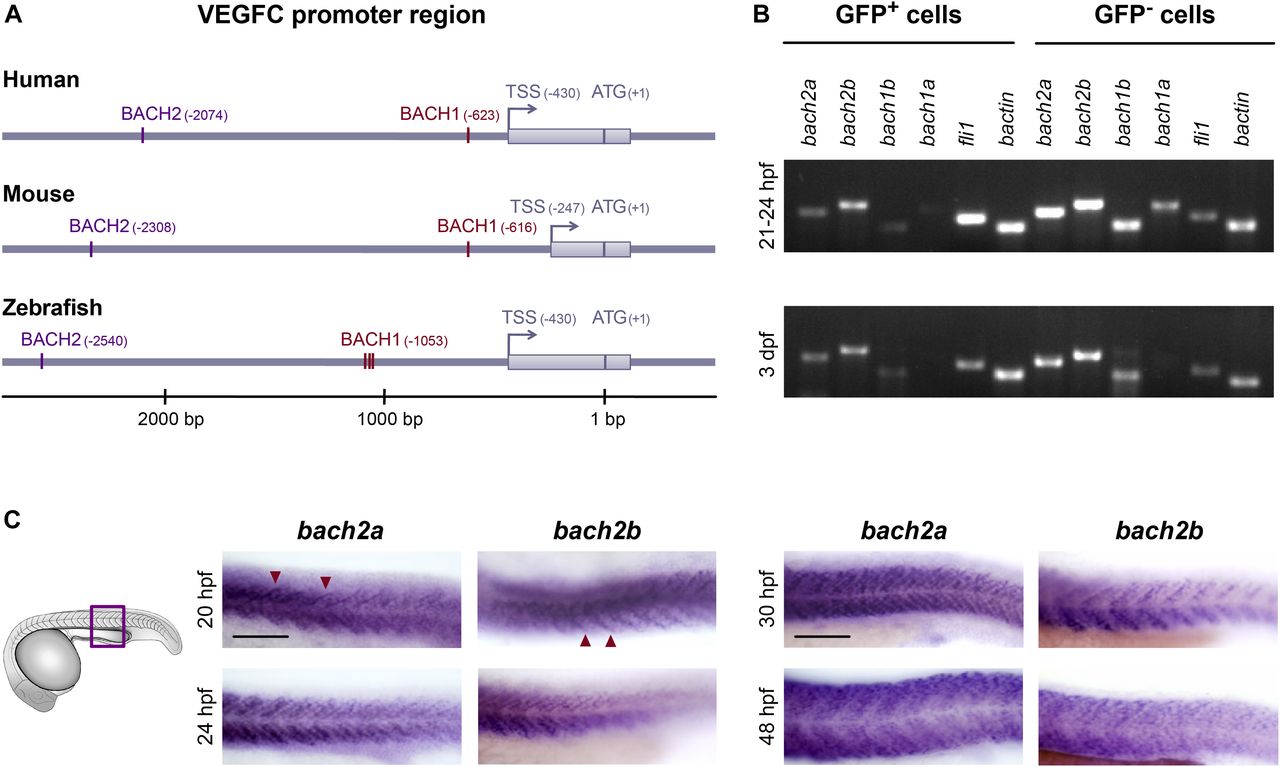
(A) BACH putative binding sites are evolutionarily conserved in the VEGFC promoter region. Numbering is from the ATG (translation initiation) because of the difference in length of the mouse’s 5′ UTR. An arrow indicates transcription start site (TSS), and the location of the first exon is marked as a gray rectangle. The location of the BACH sites is as predicted by Genomatix Genome Analyzer MatInspector: Human (NM_005429.5 TSS at hg19, chr4:177713899 on the minus strand); Mouse (NM_009506.2 TSS at mm9, chr8:54077532 on the plus strand); and Zebrafish (NM_205734.1 TSS at Zv9, chr1:39270725 on the minus strand). (B) Semi-quantitative RT-PCR analysis of the indicated genes (bactin-β actin) in enriched GFP+ cells isolated by FACS from Tg(fli1:EGFP)y1 embryos at two developmental time points, 21–24 hpf and 3 dpf (two independent experiments for each time point). (C) A lateral view of the trunk region of a wild-type zebrafish embryo at 20, 24, 30, and 48 hpf, detected with a specific bach2a or bach2b anti-sense mRNA probe. A red arrowhead indicates somite boundaries. Scale bar, 100 μm.
The BACH family of transcription factors comprises two members, BACH1 and BACH2. In mammals, BACH1 is expressed ubiquitously and has been shown to act either as an activator or repressor of transcription and to be involved in oxidative stress, metabolism, cell transformation, neurodegenerative diseases, tumor expansion, and metastatic spread (Watari et al, 2008; Warnatz et al, 2011; Nakanome et al, 2013; Igarashi & Watanabe-Matsui, 2014; Lee et al, 2014; Zhou et al, 2016; Lee et al, 2019; Lignitto et al, 2019; Wiel et al, 2019). BACH2 is a transcriptional repressor crucial for the terminal differentiation and maturation of both T and B lymphocytes (Sidwell & Kallies, 2016), and its loss is associated with severe autoimmune diseases. BACH proteins are highly conserved in vertebrates, particularly in the functional (BTB and bZip) domains and the regions immediately surrounding them (Igarashi & Watanabe-Matsui, 2014). Zebrafish have four bach genes with homology to mammalian BACH: bach1a, bach1b, bach2a, and bach2b (Zhang et al, 2014; Fuse et al, 2015; Luo et al, 2016).
As a first step, we evaluated the expression pattern of the different bach transcripts in ECs isolated by FACS from Tg(fli1:EGFP)y1 (Lawson & Weinstein, 2002) zebrafish embryos (Covassin et al, 2006a). We found the transcripts of bach2a and bach2b to be more abundant than bach1b, whereas bach1a is barely detectable in the GFP-positive (GFP+) cell population, at either 21–24 hours post-fertilization (hpf) or 3 days post-fertilization (dpf) (Fig 1B). Consequently, we focused our studies on bach2a and bach2b, as they are the major paralogs expressed in the endothelium. To assess the spatial and temporal expression patterns of bach2a and bach2b during zebrafish embryogenesis, we performed whole-mount in situ hybridization. Abundant expression of bach2a and bach2b was apparent in the somites and somite boundaries at 20 hpf and up to 48 hpf (Fig 1C). Furthermore, we detected strong enrichment of bach2a transcripts in myotomes and in several areas of the central nervous system, including the hindbrain, midbrain–hindbrain boundary, midbrain, and forebrain (Fig S1).

(A, B, C, D, E) Embryos at 20 (A), 24 (B), 30 (C, D), and 48 (E) hpf were examined by whole-mount in situ hybridization with an antisense bach2a riboprobe. Abbreviations: FB, forebrain; HB, hindbrain; M, myotomes; MB, midbrain; MHB, midbrain hindbrain boundary; OT, optic tectum; rHB, rostral hindbrain; S, somite; SB, somite-boundaries; TE, telencephalon. Scale bar, 500 μm.
bach2a is essential for developmental angiogenesis and lymphangiogenesis
We then assessed the contribution of BACH to angiogenesis and lymphangiogenesis during embryonic development and, specifically, investigated their putative role as regulators of vegfc expression and function during zebrafish vasculature formation. We used antisense morpholino oligonucleotides (MOs) to knockdown the expression of the two bach2 paralogs and then surveyed the phenotypic changes at different stages of development. At 30 hpf, clear defects were observed in the primordial hindbrain channel (PHBC) in bach2a MO-injected Tg(fli1:EGFP)y1 zebrafish embryos (Fig 2A and B). Similar phenotypes were detected in vegfc morphants (Fig 2A and B), in line with previous reports demonstrating impaired PHBC formation following the blocking of Vegfc-activated Vegfr3 signaling (Covassin et al, 2006b; Hogan et al, 2009b; Villefranc et al, 2013; Schuermann et al, 2015; Shin et al, 2016). In contrast to bach2a down-regulation, no defects were detected in Tg(fli1:EGFP)y1 embryos either injected with different concentrations of bach2b MOs (up to 10 ng per embryo, Fig 2A and C) or subjected to a bach2b CRISPR gRNA (bach2b gRNA, Figs S2A and B and 2A and B). The vascular abnormalities detected upon bach2a and vegfc knockdown were accompanied by pericardial and body edema, as well as reduced blood flow (Fig S3), resembling the phenotypes observed in collagen and calcium-binding EGF domain-1 (ccbe1) (Hogan et al, 2009a) and vegfc (Karkkainen et al, 2004; Shin et al, 2016) mutants. In contrast, no morphological abnormalities were observed after bach2b knockdown (Fig S4A–C).
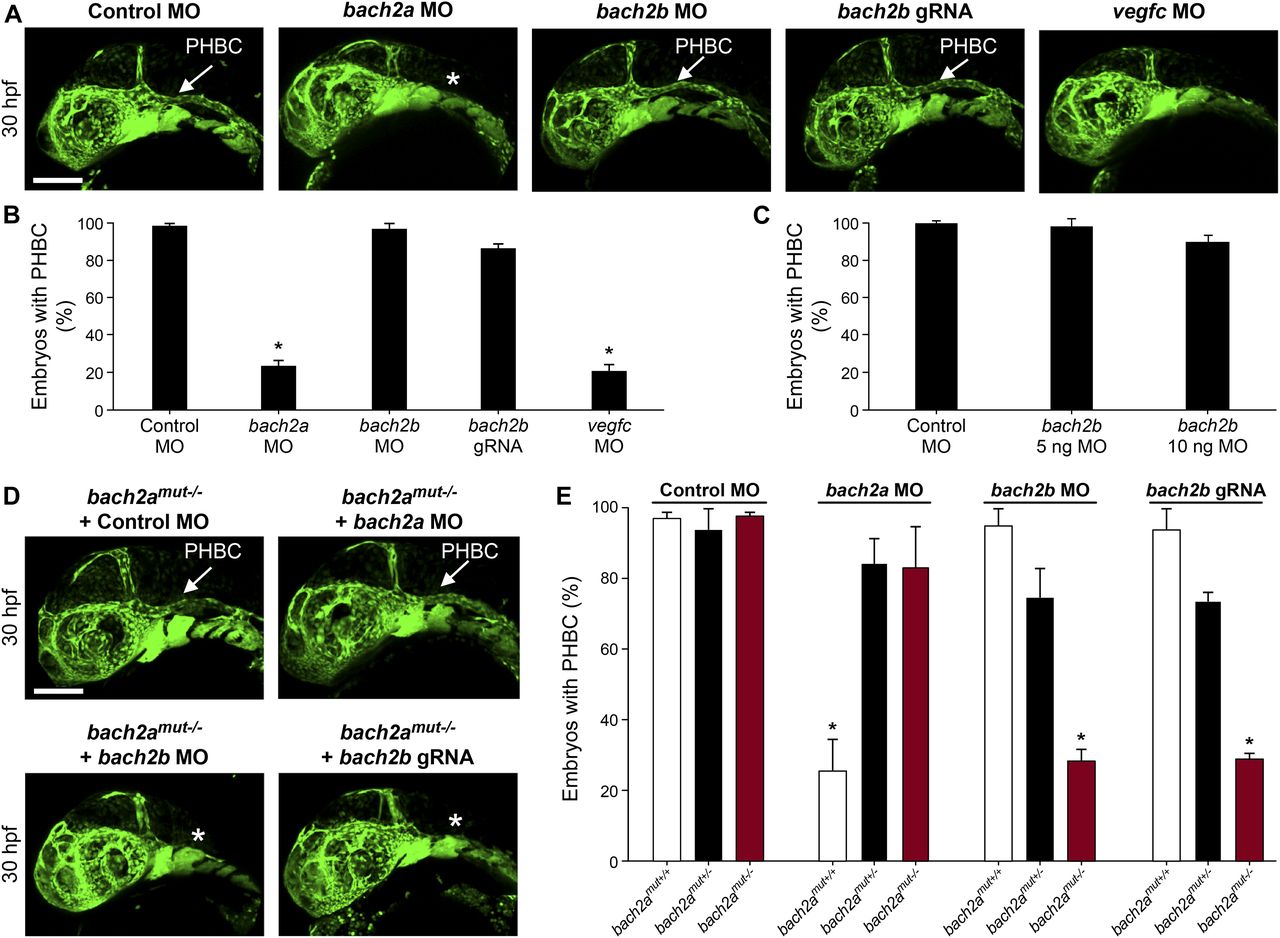
(A) Confocal images of the primordial hindbrain channel (PHBC, white arrow) of 30-hpf Tg(fli1:EGFP)y1 embryos injected with control MO (10 ng), bach2a MO (3.75 ng), bach2b MO (3.75 ng), bach2b gRNA (125 ng), or vegfc MO (10 ng). Asterisk indicates the absence of PHBC. (B) Percentage of 30-hpf Tg(fli1:EGFP)y1 embryos with intact PHBC formation after injection with control MO (10 ng, nControl MO = 68), bach2a MO (3.75 ng, nbach2a MO = 107; *P < 0.0001), bach2b MO (3.75 ng, nbach2b MO = 48), bach2b gRNA (125 ng, nbach2b gRNA = 42), or vegfc MO (10 ng, nvegfc MO = 35; *P < 0.0001). Error bars, mean ± SEM. (C) Percentage of 30-hpf Tg(fli1:EGFP)y1 embryos with intact PHBC formation after injection with control MO (10 ng, nControl MO = 24) or an increased concentration of bach2b MO (5 ng, nbach2b MO 5ng = 24) or (10 ng, nbach2b MO 10ng = 24). Error bars, mean ± SEM; P > 0.99999. (D) Confocal projection at 30 hpf Tg(fli1:EGFP)y1 of homozygous bach2a mutants (bach2amut−/−) from F2 bach2amut+/− incross. White arrow points at an intact PHBC detected in embryos injected with control MO (10 ng, bach2amut−/− + Control MO) and bach2a MO (3.75 ng, bach2amut−/− + bach2a MO). Asterisk indicates defects in PHBC development after injection with bach2b MO (3.75 ng, bach2amut−/− + bach2b MO) or bach2b gRNA (125 ng, bach2amut−/− + bach2b gRNA). Scale bar, 100 μm. (E) Percentage of randomly selected bach2a+/− F2 incross progeny at 30 hpf with an intact PHBC formation injected with control MO (10 ng, nbach2amut + Control MO = 50; P > 0.99999), bach2a MO (3.75 ng, nbach2amut + bach2a MO = 75; *P < 0.0002), bach2b MO (3.75 ng, nbach2amut + bach2a MO = 55; *P < 0.012) or bach2b gRNA (125 ng, nbach2amut + bach2b gRNA = 137; *P < 0.001). After genotyping, offspring followed the expected Mendelian ratios of inheritance. Error bars, mean ± SEM. Kruskal–Wallis test in panels (B, C, E).

(A) Schematic illustration of the bach2b locus (XM_005160548.3) and the gRNA targeting exon 4. The protospacer-adjacent motif (PAM) sequence is shown in red letters and the cleavage site is indicated by a red arrowhead. (B) Agarose gel electrophoresis of PCR products amplified from 24-hpf wild-type (WT) or bach2b gRNA (125 ng, bach2bmut)–injected Tg(fli1:EGFP)y1 embryos using primer sets which flank the gRNA cut sites. Arrows indicate the lengths of the resultant DNA fragments. (C) Schematic diagram of the bach2a locus (GRCz10, chr17:15666119–15734205). The CRISPR guide was designed to target exon 2, with the cleavage site (red arrowhead) located 115 bp downstream of the ATG. (D) Representative genotyping PCR of a wild-type (WT) and a CRIPSR/Cas9–mediated homozygous bach2a knockout (bach2amut−/−) Tg(fli1:EGFP)y1 embryo using primer sets, which flank the gRNA cut sites. Arrows indicate the lengths of the resultant DNA fragments. (E) PCR-sequencing chromatograph of the bach2a locus in a Tg(fli1:EGFP)y1 embryo (WT) aligned with a homozygous bach2a knockout (bach2amut−/−). CRIPSR/Cas9–mediated genome modification at the zebrafish bach2a locus leads to the deletion of five base-pairs, which causes a frame shift in the protein sequence (translation shown below the DNA sequence). A stop codon (indicated by a dot in the translation) created 17 amino acids downstream of the deletion site, leading to the formation of a 57-amino acid–long, truncated protein. (F) Quantitative RT-PCR validation of bach2b expression conducted for wild-type (bach2amut+/+) and homozygous (bach2amut−/−) 6-dpf siblings obtained from F2 bach2a intercross. Error bars; mean ± SEM; P = 0.2466; Wilcoxon rank sum test (two independent experiments consisting of 20 single larvae in each group). (G) Semi-quantitative RT-PCR analysis of bach2a expression in wild-type (bach2amut+/+) and homozygous (bach2amut−/−) 6-dpf siblings obtained from F2 bach2a heterozygous intercross.
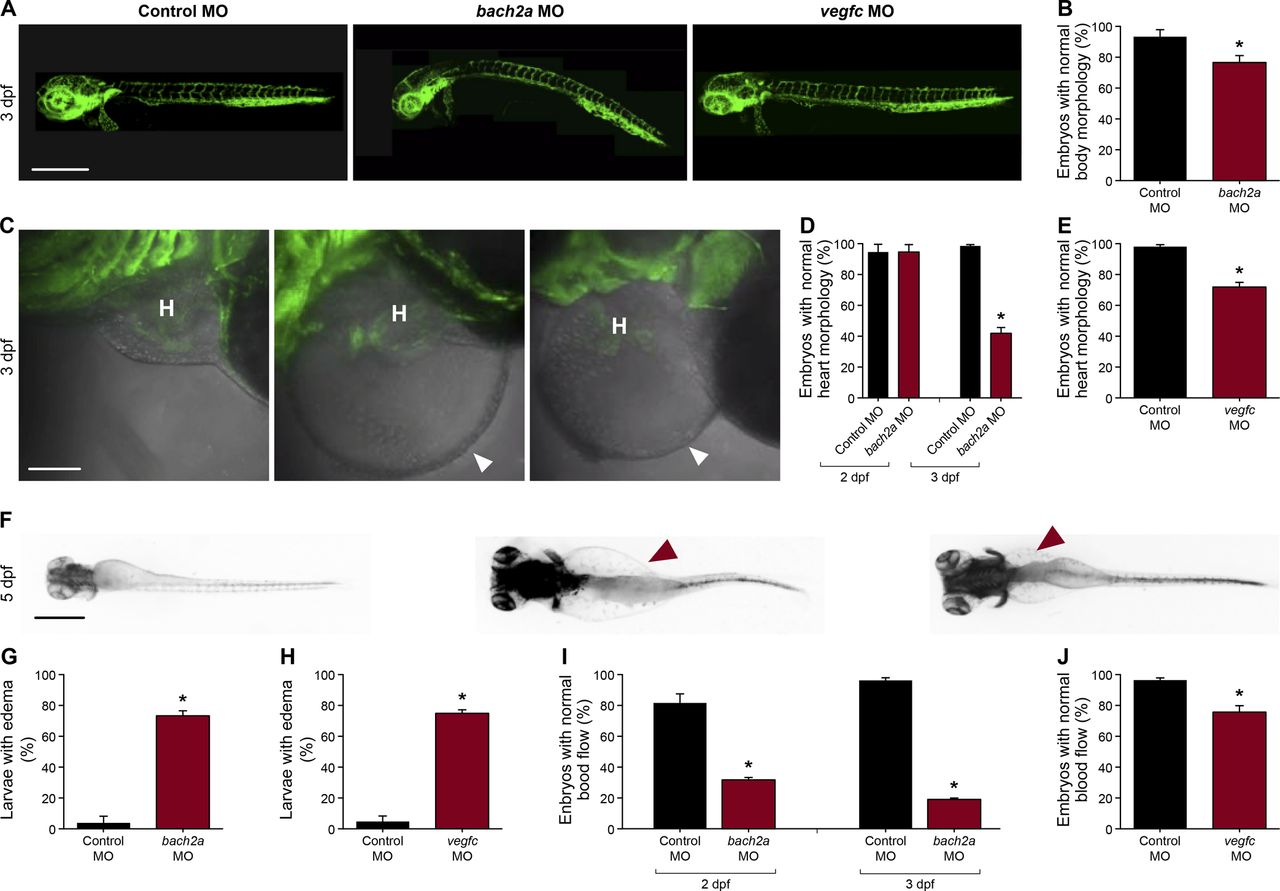
(A) Lateral-view confocal micrographs of full-body, 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos after injection with control (10 ng, Control MO), bach2a (3.75 ng, bach2a MO), or vegfc (10 ng, vegfc MO) morpholino oligonucleotide, showing body curvature of bach2a morphants. Scale bar, 500 μm. (B) Percentage of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos with a normal morphology (no body curvature) after injection with control (10 ng, Control MO) or bach2a MO (3.75 ng, nControl MO = 46; nbach2a MO = 47 *P ≤ 0.0005). (C) Confocal images of the heart morphology of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos after injection with control, bach2a, or vegfc MO. H marks the heart and a white arrowhead, a pericardial edema. Scale bar, 500 μm. (D) Quantification of 2- and 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos with a normal heart morphology after injection with control MO (10 ng, Control MO) or bach2a MO (3.75 ng, 2 dpf: nControl MO = 16; nbach2a MO = 19; 3 dpf: nControl MO = 47; nbach2a MO = 58; *P < 0.0001). (E) Percentage of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos with a normal heart morphology after injection with control MO (10 ng) or vegfc MO (10 ng, nControl MO = 54; nvegfc MO = 41; *P < 0.0001). (F) Dorsal view of 5-dpf Tg(fli1:EGFP)y1 zebrafish larvae after injection with control MO, bach2a MO, or vegfc MO. Red arrowhead indicates edema. Scale bar, 100 μm. (G) Percentage of 5-dpf Tg(fli1:EGFP)y1 zebrafish larvae with edema after injection with control MO (10 ng) or bach2a MO (3.75 ng, nControl MO-10ng = 22; nbach2a MO = 28; *P < 0.05). (H) Quantification of 5-dpf Tg(fli1:EGFP)y1 zebrafish larvae with edema after injection with control MO (10 ng) or vegfc MO (10 ng, nControl MO = 22; nvegfc MO = 53; *P < 0.0001). (I) Percentage of 2- and 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos with a normal blood flow rate after injection with control MO (10 ng) or bach2a MO (3.75, 2 dpf: nControl MO = 14; nbach2a MO = 19; 3 dpf: nControl MO = 47; nbach2a MO = 58; *P < 0.05). (J) Percentage of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos with a normal blood flow rate after injection with control MO (10 ng) or vegfc MO (10 ng: nControl MO = 54; nvegfc MO = 41; *P < 0.004). (B, D, E, G, H, I, J) Wilcoxon rank sum test in panels (B, D, E, G, H, I, J).

(A) Percentage of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos after injection with control MO (10 ng) or bach2b MO (3.75, 5, or 10 ng) showing normal heart morphology (nControl MO = 18; nbach2b MO-3.75ng = 30; nbach2b MO-5ng = 30; nbach2b MO-10ng = 10; P ≥ 0.3878). (B) Quantification of 5-dpf Tg(fli1:EGFP)y1 zebrafish larvae with edema after injection with control MO (10 ng) or bach2b MO (3.75, 5, or 10 ng) (nControl MO = 18; nbach2b MO-3.75ng = 30; nbach2b MO-5ng = 30; nbach2b MO-10ng = 10; P > 0.9999). (C) Percentage of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos with a normal blood flow rate after injection with control MO (10 ng) or bach2b MO (3.75 and 5 ng) (nControl MO = 18; nbach2b MO-3.75ng = 30; nbach2b MO-5ng = 30; P > 0.9999). (A, B, C) Kruskal–Wallis test in panels (A, B, C).
To confirm the specificity of the bach2a morphant phenotype, we generated bach2a mutants using CRISPR/Cas9–mediated gene editing (Fig S2C–E). To our surprise, homozygous bach2a mutants displayed no PHBC defects (bach2amut−/−, Fig 2D and E). We, therefore, hypothesized that genetic compensation (Rossi et al, 2015; El-Brolosy & Stainier, 2017; El-Brolosy et al, 2019) through activation of bach2b could potentially account for the absence of angiogenic phenotypes in bach2a mutants. To address this possibility, we mated bach2a+/− carriers and injected their progeny with either a sub-dose of bach2b MO or with bach2b gRNA that does not induce vascular malformations in WT embryos (Fig 2A–C). Interestingly, PHBC formation defects were identified in ∼25% of the embryos, which upon genotyping were found to carry the bach2a mutation (bach2amut−/− + bach2b MO and bach2amut−/− + bach2b gRNA, Fig 2D and E), as opposed to their wild-type siblings (bach2amut+/+ + bach2b MO and bach2amut+/+ + bach2b gRNA, Fig 2E). Conversely, after bach2a MO injection, bach2a wild-type siblings displayed PHBC formation defects, whereas homozygous mutants appeared normal (Fig 2D and E), suggesting that the bach2amut−/− mutants were less sensitive than their bach2amut+/+ siblings to bach2a MO injections, and further confirming the specificity of both mutants and morphants. To ascertain whether the compensation mechanism involves up-regulation of the paralogous Bach2b gene, we assessed its expression in wild-type and bach2a mutants, by quantitative real-time-PCR. Surprisingly, although no significant differences in the levels of bach2b mRNA were detected (Fig S2F), we observed a slight elevation in bach2a mRNA levels in homozygous mutants (Fig S2G), suggesting a non-transcriptional mechanism. The underlying molecular mechanisms by which paralogous transcription factors compensate for each other’s loss-of-function are relatively unexplored. A wide range of mechanisms may provoke robustness involving posttranscriptional or posttranslational regulation and pleiotropic effects. In addition, compensation may take place at the level of protein–protein interactions, whereby paralogs replace each other with respect to their binding partners through ancestrally preserved binding ability. Thus, we speculate that other compensatory mechanisms (not transcriptional), yet unknown, may contribute to the compensation mechanism controlling bach robustness.
We then asked whether bach2a is also involved in lymphatic vessel development. To answer this question, we analyzed the effects of bach2a down-regulation on the formation of parachordal cells (PACs), the building blocks of the thoracic duct (TD), and the trunk lymphatic system in zebrafish (Yaniv et al, 2006; Nicenboim et al, 2015). A significantly reduced number of PAC-containing segments was detected in bach2a MO–injected Tg(fli1:EGFP)y1 embryos at 3 dpf (Fig 3A and B), recapitulating the vegfc MO induced phenotype (Fig 3A and C) and the previously reported phenotype of vegfc mutants (Villefranc et al, 2013). In addition, bach2a morphants failed to express the lymphatic endothelial marker lyve1, but the expression of vegfc receptor flt4 remained unchanged (Fig S5). To rule out the possibility that the observed phenotypes are a consequence of developmental delay, we assessed the formation of the TD at 4 dpf. MO-mediated down-regulation of bach2a resulted in a significant decrease in TD formation (76%, Fig 4A and B) as compared with control MO–injected siblings. Similarly, 87% of vegfc morphants were devoid of a TD (Fig 4A and C), as previously reported (Villefranc et al, 2013; Shin et al, 2016). In contrast to bach2a morphants, bach2b MO (Figs 3A and D and 4A and D) and bach2b gRNA-injected Tg(fli1:EGFP)y1 embryos (Figs 3A and E and 4A and E) exhibited no lymphatic defects. In line with the possible genetic compensation underlying the absence of a clear angiogenic phenotype at early developmental stages (Fig 2D and E), injection of either a sub-dose of bach2b MO or bach2b gRNA into the progeny of bach2a+/− intercross leads 23% of embryos to display significant lymphangiogenic defects, which were subsequently identified by genotyping as homozygous mutants (bach2amut−/− + bach2b MO or bach2amut−/− + bach2b gRNA) (Figs 3F and G and 4F and G). In contrast, control MO–injected bach2amut−/− appeared normal (Figs 3F and G and 4F and G). Likewise, no lymphatic phenotype was detected in bach2amut−/− embryos upon injection of bach2a MO (Figs 3F and G and 4F and G), indicating that bach2a MO has minimal off-target effects. Finally, cardiac and body edema, characteristic of lymphatic-related defects, were detected in bach2a mutants after bach2b MO or bach2b gRNA injection, but not upon bach2a MO administration (Fig S6). Remarkably, the phenotypic defects resulting from bach2a down-regulation were more pronounced than those caused by the loss of vegfc, possibly due to the involvement of bach2a in additional signaling pathways (Zhou et al, 2016).

(A, B) Confocal projection of the trunk of 3-dpf Tg(fli1:EGFP)y1 embryos showing PACs (white arrow) in control MO, bach2b, or bach2b gRNA-injected embryos but not after injection with bach2a or vegfc MO (white asterisk). Scale bar, 100 μm. (B) Number of PAC-containing segments (mean ± SEM) in 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos injected with control MO (10 ng) or bach2a MO (3.75 ng, nControl MO = 46; bach2a MO, nbach2a MO = 53; *P < 0.001). Error bars, mean ± SEM. (C) Number of PAC-containing segments in vegfc MO-injected morphants (10 ng, nControl MO = 53, nvegfc MO = 41; *P < 0.0001). Error bars, mean ± SEM. (D) Number of PAC-containing segments in 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos injected with indicated bach2b MO concentrations (3.75, 5, or 10 ng, nControl MO = 18; nbach2b MO-3.75ng = 30; nbach2b MO-5ng = 30; nbach2b MO-10ng = 10; P ≥ 0.2819). Error bars, mean ± SEM. (E) Quantification of PAC-containing segments in embryos injected with bach2b gRNA (125 ng, nControl MO = 35 nbach2b gRNA = 42; P = 0.0615). Error bars, mean ± SEM. (F) Confocal projection of the trunk region showing PAC-containing segments in Tg(fli1:EGFP)y1-homozygous bach2a mutants (bach2amut−/−) from F2 bach2amut+/− incross. An asterisk indicates the absence of PACs in bach2amut−/− embryos injected with bach2b MO (bach2amut−/− + bach2b MO) or bach2b gRNA (bach2amut−/− + bach2b gRNA) and a white arrow, their presence. Scale bar, 100 μm. (G) Number of PAC-containing segments (mean ± SEM) in 3-dpf embryos randomly selected from bach2a+/− F2 incross progeny injected with control MO (10 ng, nbach2amut + Control MO = 50; P = 0.5514), bach2a MO (3.75 ng, nbach2amut + bach2a MO = 75; *P < 0.0001), bach2b MO (3.75 ng, nbach2amut + bach2b MO = 55; *P and **P < 0.0222), or bach2b gRNA (125 ng, nbach2amut + bach2b gRNA = 137; *P and **P < 0.0001). After genotyping, offspring followed the expected Mendelian ratios of inheritance. (B, C, D, E, G) Wilcoxon rank sum test in panels (B, C, E) and Kruskal–Wallis test in panels (D, G).

(A) Drawing of zebrafish embryo. (B, C) Purple box indicates region imaged in panels (B, C). (B) Whole-mount in situ hybridization comparing the expression pattern (red arrowhead) of lyve1 in 24-hpf wild-type zebrafish embryos after injection with control or bach2a MO (red asterisk indicates absence of expression). (C) Whole-mount in situ hybridization using flt4 riboprobe of 24-hpf, wild-type zebrafish embryos after injection with control or bach2a MO. Scale bar, 100 μm.

(A) Confocal images of the TD in 4-dpf control MO- (10 ng), bach2a MO-(3.75 ng), vegfc MO- (10 ng), bach2b MO- (10 ng), or bach2a gRNA-injected Tg(fli1:EGFP)y1 larvae. A white arrow indicates the presence of a TD and a white asterisk, its absence. Scale bar, 20 μm. (B) Percentage of 4-dpf Tg(fli1:EGFP)y1 larvae with an intact TD after injection with control MO (10 ng) or bach2a MO (3.75 ng, nControl MO = 46; nbach2a MO = 52). Error bars, mean ± SEM; *P < 0.0001. (C) Percentage of larvae with a TD after injection with vegfc MO (10 ng, nControl MO = 46; nvegfc MO = 52). Error bars, mean ± SEM; *P < 0.0001. (D) Quantification of bach2b morphants with an intact TD after injection with indicated bach2b MO concentrations (3.75, 5, or 10 ng, nControl MO = 18; nbach2b MO-3.75ng = 30; nbach2b MO-5ng = 30; and nbach2b MO-10ng = 10; P ≥ 0.1356). Error bars, mean ± SEM. (E) Percentage of TD-containing larvae injected with bach2b gRNA (125 ng, nControl MO = 35 nbach2b gRNA = 42; P = 0.5461). Error bars, mean ± SEM. (F) Confocal images of a TD (white arrow) in 4-dpf Tg(fli1:EGFP)y1: homozygous bach2a mutants (bach2amut−/−) derived from bach2amut+/− F2 incross. An asterisk indicates absence of a TD and a white arrow, its presence. Scale bar, 20 μm. (G) Analysis of Tg(fli1:EGFP)y1 4-dpf progeny obtained from F2 bach2a heterozygous intercross. Random selection from the pool of siblings injected at the one-cell stage with control MO (10 ng, nbachmut + Control MO = 50), bach2a MO (3.75 ng, nbach2amut + bach2a MO = 75), bach2b MO (3.75 ng, nbach2amut + bach2b MO = 55), or bach2b gRNA (125 ng, nbach2amut + bach2b gRNA = 137) was found to maintain, after genotyping, the expected Mendelian ratios of inheritance. Error bars, mean ± SEM; *P < 0.0003. (B, C, D, E, G) Wilcoxon rank sum test in panels (B, C, E) and Kruskal–Wallis test in panels (D, G).

(A) Fluorescent images of homozygous bach2a (bach2amut−/−) siblings obtained from F2 bach2a heterozygous intercross injected at the one-cell stage with control (10 ng, Control MO) or bach2b MO (3.75 ng). Scale bar, 500 μm. (A, B) Bright-field images corresponding to the larvae presented in panel (A). Red arrowhead indicates cardiac edema. Scale bar, 500 μm. (C) Lateral view of 6-dpf homozygous bach2a (bach2amut−/−) siblings obtained from F2 bach2a heterozygous intercross injected with control (10 ng, Control MO), bach2a (3.75 ng, bach2a MO), bach2b (3.75 ng, bach2b MO) morpholino, or bach2b gRNA (125 ng, bach2b gRNA). Red arrowhead indicates cardiac edema, whereas body edema is indicated by a black arrow. Scale bar, 500 μm.
Taken together, these findings demonstrate that knockdown of bach2a through MO injection results in the formation of defective blood and lymphatic vascular plexuses, despite the presence of the bach2b paralog, indicating that bach2a is a major contributor to this phenotype. However, knockout of bach2a expression (in mutants) triggers a functional compensation by bach2b, suggesting that the two bach2 genes may share partially overlapping functions, which allows them to compensate for each other’s loss during blood and lymph vessel development. Accordingly, it was suggested that BACH1 and BACH2 act in a complementary manner to maintain normal alveolar macrophage function and surfactant homeostasis in the lung (Ebina-Shibuya et al, 2016). Similar phenotypic differences between mutants and transient knockdown animals were observed in various model systems, some of which were attributed to genetic compensation (Rossi et al, 2015; El-Brolosy & Stainier, 2017; El-Brolosy et al, 2019).
BACH1 promotes angiogenesis and lymphangiogenesis during tumor expansion in mouse models
In light of the perception that cancers frequently reactivate embryonic developmental signaling cascades to promote their expansion and aggressiveness, resulting in metastasis and poor patient outcome, we decided to evaluate the contribution of BACH to vascular remodeling during tumor progression. Emerging evidence point to BACH1, a ubiquitously expressed protein, as a tumor-promoting factor that acts via multiple intracellular signaling cascades (Alvarez & Woolf, 2011; Yun et al, 2011). Recently, higher levels of BACH1 were found to be associated with poor prognosis in human ovarian cancer (Han et al, 2019) and to promote lung cancer metastasis (Lignitto et al, 2019; Wiel et al, 2019). BACH1 has been established as a major regulator of breast cancer bone metastasis (Liang et al, 2012) and was postulated as a potential novel therapeutic candidate for cancer treatment (Davudian et al, 2016a; Lee et al, 2019). To assess whether enhanced expression of BACH1 in tumor cells can stimulate blood and lymphatic vessel expansion during tumor progression as well as metastatic spread, we analyzed various ovarian and lung mouse tumors (Figs 5 and S7). Inoculation of human ovarian clear cell carcinoma ES2 cells into the peritoneal cavity of immune-deficient CD-1 nude female mice has been established as a model of metastatic ovarian cancer (Shaw et al, 2004). In this study, we used this model to evaluate the effects of ectopic expression of BACH1 in human ovarian ES2 carcinoma cells (Fig 5). 20 d post-injection, metastases were detected in the diaphragm of animals injected with BACH1-overexpressing ES2 cells. An increase in CD34+ tumor-associated blood vessel density was apparent in diaphragm specimens derived from mice injected with BACH1-overexpressing cells, as compared with control ES2 cells (Fig 5A and B). Similar differences in the intratumoral blood vasculature were observed after subcutaneous inoculation of either BACH1-overexpressing ES2 cells (Fig S7A–C) or Bach1-overexpressing mouse D122 Lewis Lung Carcinoma cells (Eisenbach et al, 1984) (Fig S7E–G). These results indicate that, in addition to its role in embryonic vascular development, BACH1 promotes tumor angiogenesis.
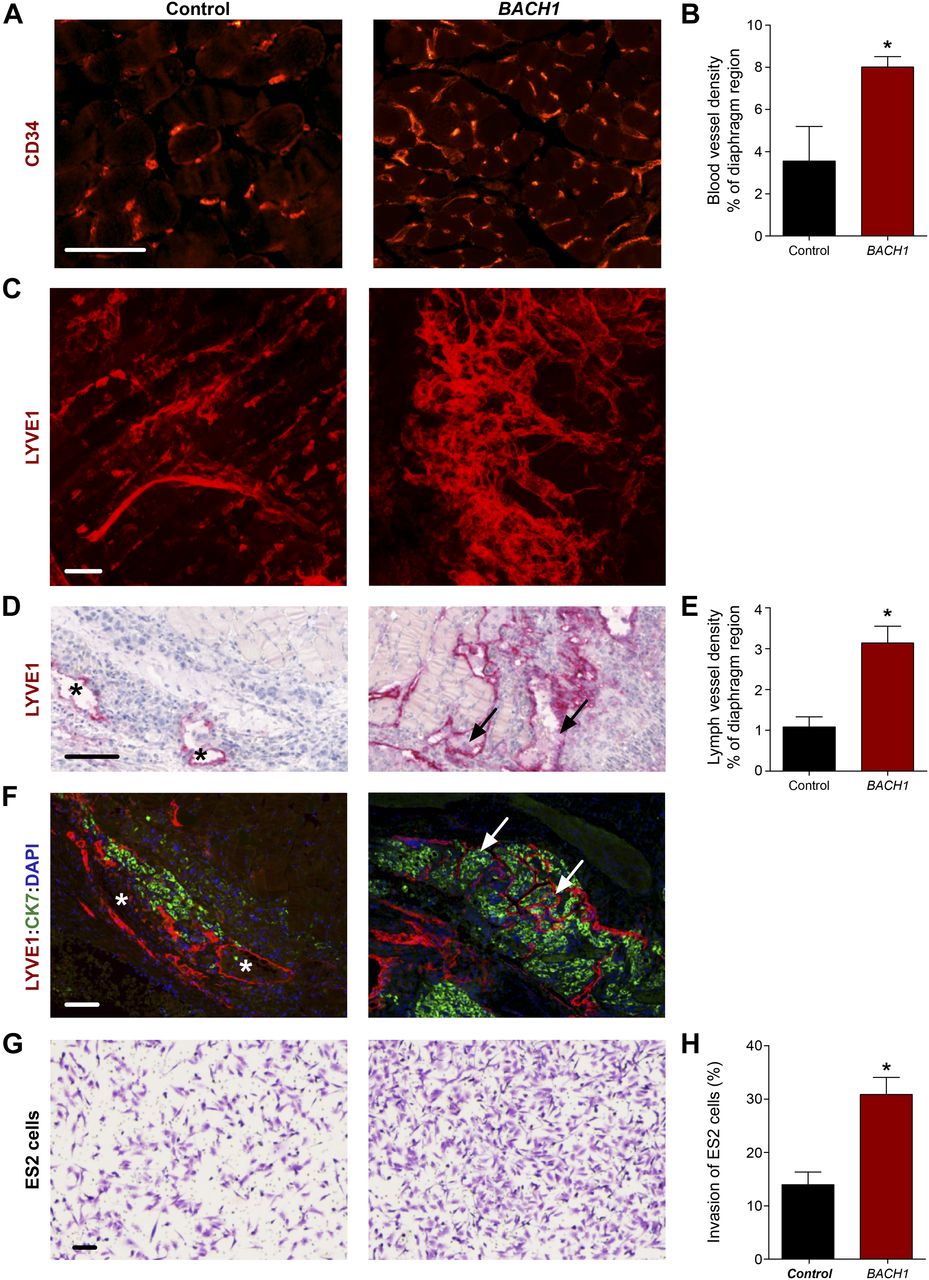
Ex vivo analysis of subcutaneous xenografts and diaphragm specimens excised from CD-1 nude female mice implanted with control (Control) or BACH1 ectopically expressing (BACH1) human ovarian clear cell carcinoma ES2 cells. (A) Immunofluorescence labeling of blood vessels using anti-CD34 antibodies in diaphragm specimens excised from mice injected intraperitoneally with control or BACH1-expressing human ES2 cells. Scale bar, 100 μm. (B) Morphometric analysis of the diaphragm relative region covered by CD34+ blood vessels. Diaphragms were excised from mice inoculated intraperitoneally with either control (Control, n = 3) or BACH1-overexpressing (BACH1, n = 7) ES2 cells (mean ± SEM; *P = 0.0304). (C) Confocal z-projection images (Z dimension 7 μm) of control and BACH1-overexpressing subcutaneous-ES2 ovarian carcinoma xenografts subjected to LYVE1 immunofluorescence staining along with a modified CLARITY technique. Images demonstrate the complexity of the lymphatic vasculature. Scale bar, 100 μm. 3D reconstructions of the stacks are available in Videos 1 and 2. (D) Lymphatic vessel immunostaining, using anti-LYVE1 antibodies, of diaphragm specimens excised from mice injected intraperitoneally with control or BACH1-overexpressing ES2 ovarian carcinoma cells. A black arrow indicates infiltration of cells into the lymphatic vessel and an asterisk, their absence. Scale bar, 100 μm. (E) Morphometric analysis of the diaphragm relative region covered by LYVE1+ lymph vessels. Diaphragms were excised from mice inoculated intraperitoneally with either control (Control, n = 6) or BACH1-overexpressing (BACH1, n = 9) ES2 cells (mean ± SEM; *P = 0.0047). (F) Immunofluorescence double staining of LYVE1+ lymphatic vessels (red) and cytokeratin 7 (CK7, green) of a 4-μm-thick specimen sectioned from paraffin-embedded diaphragm excised from mouse inoculated intraperitoneally with either control or BACH1-overexpressing ES2 cells. Nuclei were counterstained with DAPI (blue). A white arrow indicates infiltration of tumor cells into the lymphatic vessels and an asterisk, their absence. Scale bar, 100 μm. (G) Transwell Matrigel invasion assay performed in vitro with ES2 ectopically expressing BACH1 and control cells. The crystal violet dye staining images of the lower chambers are shown. Scale bar, 100 μm. (G, H) Percentage of ES2 cells that invaded through the Matrigel matrix (as in panel G) normalized to total cell number (n = 2 for each group, in duplicates; mean ± SEM; *P = 0.0209). (B, E, H) Wilcoxon rank sum test in panels (B, E, H).

Analysis of subcutaneous tumor specimens (two mouse models: human ovarian clear cell carcinoma ES2 cells in CD-1 nude mice and mouse D122 Lewis lung carcinoma cells in C57BL/6 black mice) excised from mice implanted with either ectopic overexpression of HA-BACH1 or control cells expressing an empty vector (Control). (A) Western blot detecting HA-BACH1 and β-tubulin in protein extracts from control (−) or BACH1 (+)-ectopically expressing ES2 cell. (B) Immunofluorescence labeling of blood vessels using anti-CD34 antibodies of subcutaneous tumors excised from mice inoculated with control (Control) or BACH1 (BACH1)-ectopically expressing human ES2 cells. (C) Morphometric analysis of the relative coverage of CD34+ blood vessels within the tumor area (n = 5 in each group; mean ± SEM; *P < 0.0001). (D) Immunohistochemical staining of lymphatic endothelial cells using anti–LYVE1 antibodies. Scale bar, 100 μm. (E) Western blot detecting HA-BACH1 and β-tubulin in protein extracts from control (−) or Bach1 (+)-ectopically expressing mouse D122 Lewis lung carcinoma cells. (F) Immunofluorescence labeling of blood vessels using anti-CD34 antibodies of subcutaneous tumor specimens excised from mice inoculated with control (Control) or Bach1 (Bach1)-ectopically expressing mouse D122 Lewis lung carcinoma cells. (G) Morphometric comparison between the coverage of CD34+ blood vessels within the tumor area of control (Control, n = 9) or Bach1-overexpressing (Bach1, n = 8) D122 Lewis lung cells (mean ± SEM; *P < 0.0001). (H) Immunohistochemical staining of lymphatic endothelial cells using anti-LYVE1 antibodies. (I) Quantitative RT-PCR validation of Vegfc mRNA levels in Bach1-overexpressing mouse D122 Lewis lung tumors compared with control tumor (n = 6 per group; mean ± SEM; *P < 0.05). (J) Immunohistochemical labeling of tumor sections using anti-VEGFC antibodies and counterstaining with hematoxylin (blue). Scale bar, 100 μm. (C, G, I) Wilcoxon rank sum test in panels (C, G, I).
Numerous reports suggest that, in addition to blood vessel formation, lymphangiogenesis and lymphatic vessel remodeling are pivotal events for tumor expansion and metastatic spread (Stacker et al, 2014). We, thus, examined the effects of BACH1 on the lymphatic vasculature in various mouse tumor models. To gain insight into the 3D complexity of the lymphatic network within intact tumors, we subjected ES2 subcutaneous xenografts to a modified CLARITY (Clear, Lipid-exchanged, Anatomically Rigid, Imaging-compatible, Tissue hYdrogel) technique (Hama et al, 2011; Chung et al, 2013; Oren et al, 2018), along with LYVE1 immunofluorescence staining. A robust increase in lymphatic vessel density was observed in BACH1-overexpressing tumors as compared with control-derived ES2 tumors (Fig 5C and Videos 1 and 2). Morphometric analyses indicate a significant increase in the relative area fraction occupied by LYVE1+ tumor-associated lymphatic vessels in diaphragm specimens removed from mice injected with BACH1-overexpressing ES2 cells (Fig 5D and E). In addition, enlarged lymph vessels were observed in the peritumoral region of subcutaneous tumors derived from BACH1-overexpressing ES2 and D122 cells (Fig S7D and H, respectively). Expansion of the lymph vessels in the diaphragms of BACH1-overexpressing tumors was associated with increased cell infiltration (arrow, Fig 5D). Co-immunofluorescence using antibodies directed against LYVE1 and cytokeratin 7, an antigen expressed in ES2 cells (Stimpfl et al, 1999), revealed that the metastasizing cells originated from the transplanted ES2 tumor cells (arrow, Fig 5F). This potential of BACH1 to promote tumor cell invasion was further confirmed by an in vitro invasion assay demonstrating an approximately twofold increase in the invasion abilities of BACH1-overexpressing as compared with control ES2 cells (Fig 5G and H). These data indicate that in mouse models, BACH1 remodels vascular architecture and promotes metastatic spread of tumor cells via the lymphatic vessels.
Lymphatic vessel complexity in a control tumor. A movie demonstrating the lymphatic vessel density in a control subcutaneous ES2 ovarian carcinoma xenograft subjected to LYVE1 immunofluorescence staining along with a modified CLARITY technique. Z dimension 392 μm; scale bar, 100 μm.Download video
Lymphatic vessel complexity in a BACH1-overexpressing tumor. A movie demonstrating the lymphatic vessel density in a subcutaneous BACH1-overexpressing ES2 ovarian carcinoma xenograft subjected to LYVE1 immunofluorescence staining along with a modified CLARITY technique. Z dimension: 392 μm; scale bar, 100 μm.Download video
BACH and VEGFC are functionally linked
Next, we investigated the molecular mechanisms underlying BACH activity. Because BACH proteins can directly bind to the promoter of their target genes (Ogawa et al, 2001; Warnatz et al, 2011; Yun et al, 2011), we assessed the ability of the binding sites in the VEGFC promoter region identified in silico (Figs 1A and 6A) to interact with BACH. Chromatin immunoprecipitation (ChIP) was performed on human ovarian clear cell carcinoma ES2 cells, overexpressing BACH1 tagged with a human influenza HA tag (Fig S7A). We observed that specific sequences from both proximal and distal regulatory regions were enriched in the presence of antibodies against either HA or BACH1, indicating that both sites are transcriptionally functional (Fig 6B). To investigate the functional significance of two potential BACH-binding sites, promoter–reporter constructs were engineered with deletion of the BACH-binding sites, either separately or concurrently. Disruption of the proximal site led to a 23% increase in VEGFC basal promoter activity (Fig 6C and D), supporting an inhibitory role. In contrast, deletion of the distal binding site individually or in combination with the proximal site resulted in an 86% reduction in VEGFC basal promoter activity (Fig 6C and D), suggesting a pivotal role in basal promoter functioning. In addition, overexpression of BACH1 in human ovarian ES2 carcinoma cells induces a significant increase in VEGFC expression (Fig 6E), strongly supporting a genetic and functional link between these two factors.

(A) Conservation of BACH sites in human, mouse, and zebrafish. The distal site is completely conserved. The proximal site is fully conserved between mouse and human, whereas there are three BACH sites at very close proximity in zebrafish. All proximal sites differ by one nucleotide from the consensus sequence. (B) Chromatin immunoprecipitation assay, followed by PCR measurements, was performed using primer mapping to the above human BACH proximal and distal regulatory sites and DNA precipitated with nonspecific IgG, HA-tag, or BACH1 antibodies. (C) Schematic representation of the wild-type human VEGFC promoter-driven luciferase (Luc) reporters (blue) (pVEGFCwt-Luc) and of three constructs deleted either from proximal (nt. −623 to −603, pVEGFCΔPro-Luc) or distal (nt. −2074 to −2054, pVEGFCΔDis-Luc) BACH-binding sites or a combination thereof (nt −623 to −603 and −2074 to −2054, pVEGFCΔProDis-Luc). Numbers refer to the nucleotide positions relative to ATG (translation initiation). (D) Quantification of dual–luciferase activity in human ES2 cells driven from pVEGFCwt-Luc, pVEGFCΔPro-Luc, pVEGFCΔDis-Luc, and pVEGFCΔProDis-Luc constructs. Relative luciferase activity is shown as a percentage of the pVEGFCwt-Luc value (mean ± SEM, n = 3). *P < 0.0001, Kruskal–Wallis test. (E) Immunofluorescence staining of human ES2 cells stably expressing either an empty pIRES vector (Control) or N-terminally HA-tagged BACH1 (BACH1) with antibodies directed against the HA tag (red, left panel) or against VEGFC (red, right panel). Nuclei were counterstained with DAPI (blue). Scale bar, 20 μm. TSS, transcription start site.
To investigate whether these results are recapitulated in vivo, we analyzed the effect of bach2a knockdown on vegfc expression during zebrafish development. A marked down-regulation of vegfc expression was observed in bach2a morphants (Fig 7A). This effect was specific for bach2a, as vegfc expression remained intact in bach2b MO-injected embryos (Fig S8). We then attempted to rescue the phenotypes of bach2a morphants by overexpressing vegfc mRNA. Co-injection of in vitro–transcribed vegfc mRNA and bach2a MO partially restored PHBC formation at 30 hpf (Fig 7B and C). Similarly, the number of PAC-containing segments increased by ∼50% after co-injection of bach2a MO and vegfc mRNA (Fig 7D and E) and a twofold recovery in TD formation was detected (Fig 7F and G). In addition, pericardial and body edema, as well as reduced blood flow, were partially restored after an injection of vegfc mRNA (Fig S9).
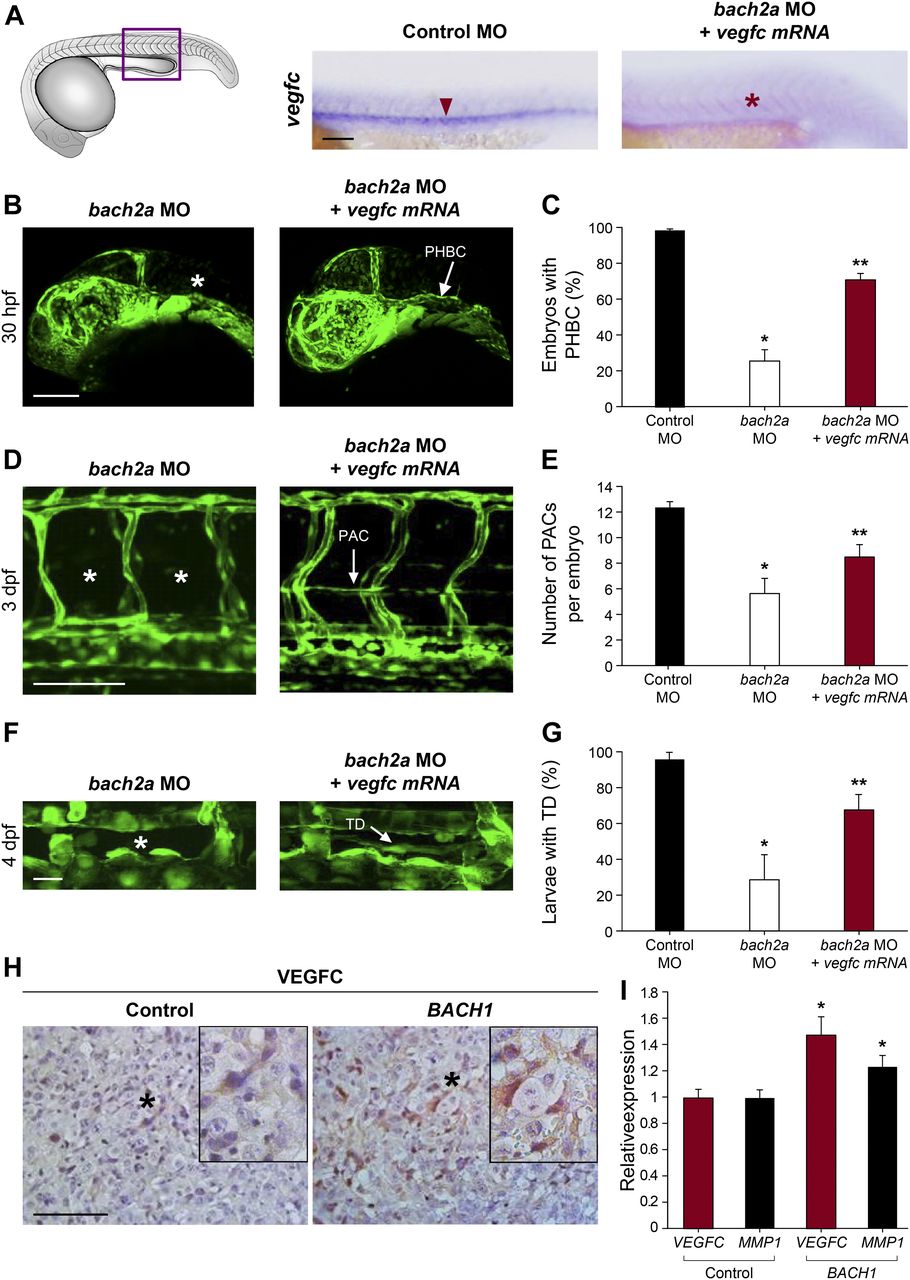
(A) Whole-mount in situ hybridization of 24-hpf, wild-type zebrafish embryos demonstrating the expression of vegfc mRNA after injection with control MO (Control MO 10 ng, red arrowhead) and the absence of its expression in embryos injected with specific MO targeting bach2a (3.75 ng, bach2a MO, red asterisk). Scale bar, 100 μm. (B) Confocal images of 30-hpf Tg(fli1:EGFP)y1 embryos co-injected with specific MOs targeting bach2a (3.75 ng) and in vitro–transcribed vegfc mRNA (800 pg, bach2a MO + vegfc mRNA) demonstrating the restoration of PHBC (white arrow). (C) Percentage of rescued PHBC defects in 30-hpf Tg(fli1:EGFP)y1 embryos after co-injection with bach2a MO (3.75 ng) and vegfc mRNA (800 pg, nControl MO = 56; nbach2a MO = 58; nbach2a MO + vegfc mRNA = 56). Error bars, mean ± SEM; * or **P < 0.0001. (D) Rescue of parachordal cell (PAC) development in 3-dpf Tg(fli1:EGFP)y1 embryos after the co-injection of bach2a MO and vegfc mRNA (bach2a MO + vegfc mRNA; PACs are indicated by a white arrow). Scale bar, 100 μm. (E) Quantification of the number of PAC-containing segments (mean ± SEM) in 3-dpf Tg(fli1:EGFP)y1 embryos after the co-injection of bach2a MO (3.75 ng) and vegfc mRNA (nControl MO = 47; nbach2a MO = 27; nbach2a MO + vegfc mRNA = 54; * or **P < 0.01). (F) Thoracic duct (TD) formation in 4-dpf Tg(fli1:EGFP)y1 embryos co-injected with bach2a MO and vegfc mRNA (bach2a MO + vegfc mRNA; TD is indicated by a white arrow). Scale bar, 20 μm. (G) Percentage of 4-dpf Tg(fli1:EGFP)y1 embryos showing normal TD after bach2a MO and vegfc mRNA injection (bach2a MO + vegfc mRNA). (nControl MO-10ng = 32; nbach2a MO = 65; nbach2a MO + vegfc mRNA = 68 (Error bars, mean ± SEM; * or **P < 0.01. (H) Immunohistochemistry labeling of control (Control) or BACH1 (BACH1) ectopically expressing ES2 ovarian carcinoma xenograft specimens using anti-VEGFC antibodies and counterstaining with hematoxylin (blue). Black asterisk localizes the region magnified in the black frame. Scale bar, 100 μm. (I) Quantitative RT-PCR measurement of VEGFC and MMP1 mRNA expression in xenograft initiated either from control or BACH1-overexpressing ES2 cells (n = 5 in each group; mean ± SEM; *P < 0.05. (C, E, G, I) Kruskal–Wallis test in panels (C, E, G) and Wilcoxon rank sum test in panel (I).
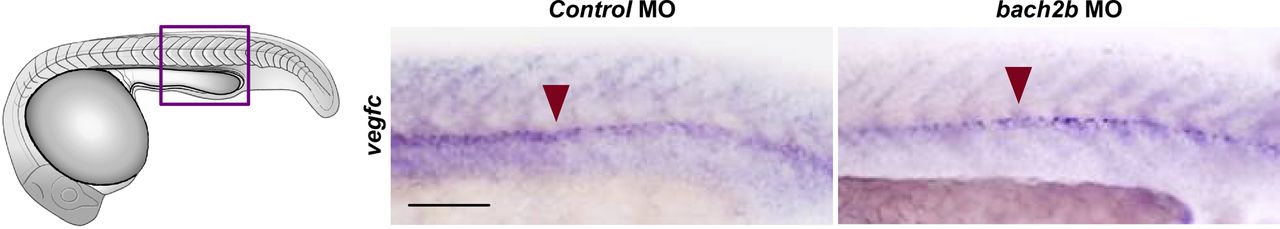
Whole-mount in situ hybridization presenting the expressions of vegfc in 24-hpf embryos injected with control or bach2b MO. Presence of expression is indicated by a red arrowhead. Scale bar, 100 μm.
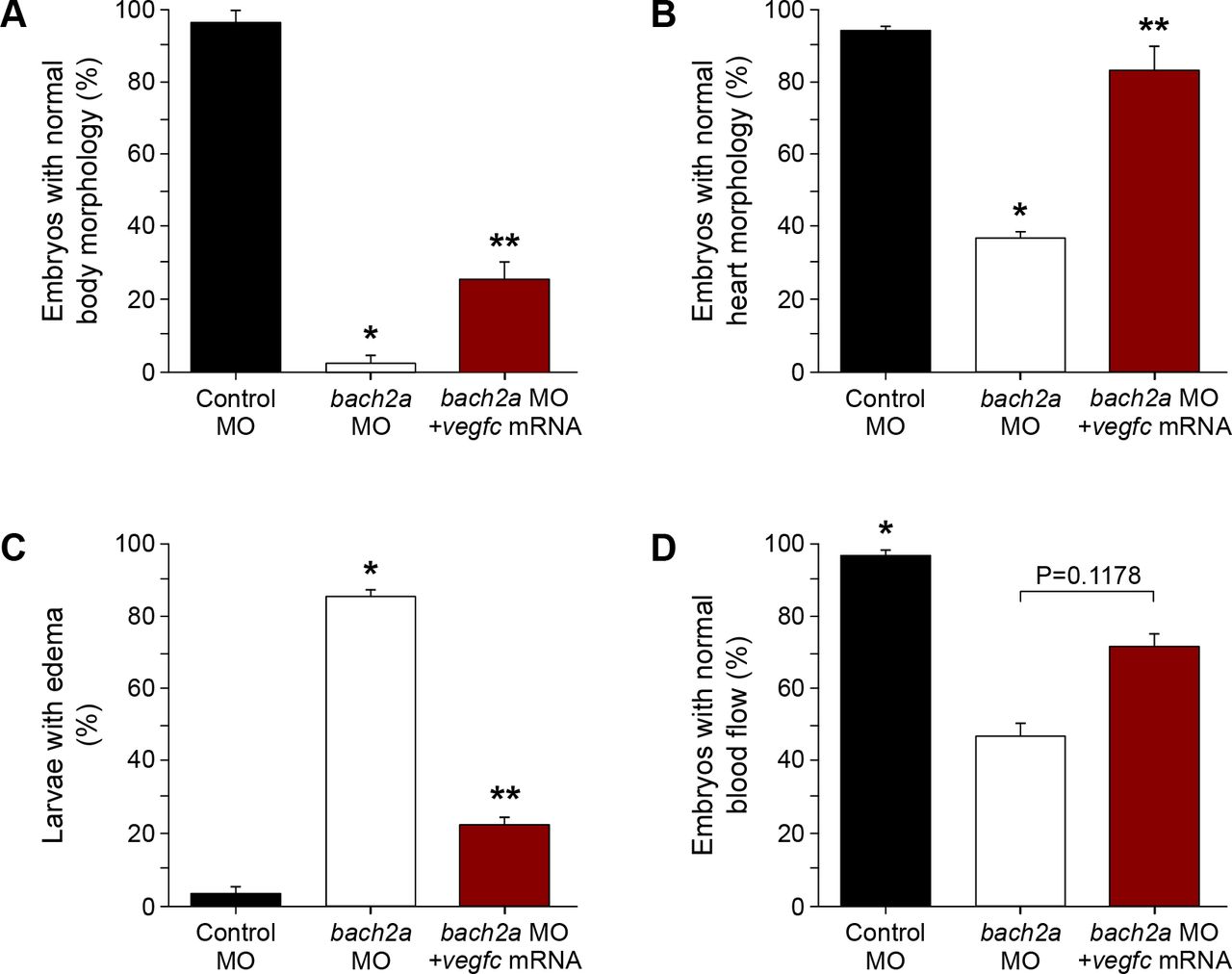
(A) Quantification of 3-dpf Tg(fli1:EGFP)y1 zebrafish embryos injected with control (10 ng), bach2a (3.75 ng) MO, or co-injection with bach2a MO (3.75 ng) together with in vitro–transcribed vegfc mRNA (800 pg) showing normal body morphology (nControl MO = 25; nbach2a MO = 41; nbach2a MO + vegfc mRNA = 43; * or **P < 0.05). (B) Percentage of normal heart morphology (3.5 dpf, b: nControl MO = 54; nbach2a MO = 28; nbach2a MO + vegfc mRNA = 33; * or **P < 0.05). (C) Percentage of rescued body edema in 5-dpf larvae (nControl MO = 25; nbach2a MO = 66; nbach2a MO + vegfc mRNA = 70; *P ≤ 0.0001 and **P < 0.05). (D) Quantification of normal blood flow rate of 3-dpf embryos (nControl MO = 54; nbach2a MO = 35; nbach2a MO + vegfc mRNA = 28, *P < 0.05). (A, B, C, D) Kruskal–Wallis test in panels (A, B, C, D).
Finally, we addressed the molecular basis of the BACH1 and VEGFC interaction during tumor progression in the various mouse tumor models. VEGFC expression was specifically elevated in BACH1-overexpressing ES2 and D122 tumors both at the protein (Figs 7H and S7J, respectively) and mRNA level (Figs 7I and S7I), with no significant changes in the mRNA levels of either VEGFA or VEGFB (Fig S10). Similarly, the mRNA expression levels of MMP1, a recognized transcription target of BACH1 (Yun et al, 2011; Liang et al, 2012), were significantly elevated in subcutaneous BACH1-overexpressing ES2 xenografts (Fig 7I). Collectively, these results highlight BACH as a novel regulator of blood and lymphatic vessel formation during both embryonic development and mouse tumor expansion, placing it upstream of VEGFC in these cascades.

(A) Quantitative RT-PCR measurement of VEGFA expression conducted on xenograft initiated either from control or BACH1-overexpressing ES2 cell (n = 5 in each group; mean ± SEM; P = 0.6857). (B) Quantitative RT-PCR validation of VEGFB expression conducted either on control or BACH1-overexpressing ES2 xenografts (n = 5 in each group; mean ± SEM; P = 0.0556). (C) Quantitative RT-PCR measurement of Vegfa mRNA levels in Bach1-overexpressing mouse D122 Lewis lung tumors compared with control tumor (n = 6 per group; mean ± SEM; P = 0.2500). (D) Quantitative RT-PCR validation of Vegfb mRNA levels in Bach1-overexpressing mouse D122 Lewis lung tumors compared with control tumor (n = 6 per group; mean ± SEM; P > 0.9999). (A, B, C, D) Wilcoxon rank sum test in panels (A, B, C, D).
Expression of BACH1 and VEGFC correlates during human cancer progression
Metastatic spread of cancer cells from primary solid tumors to sentinel lymph nodes and distant tissues and organs is one of the hallmarks of malignant neoplasms (Valastyan & Weinberg, 2011), responsible for most human cancer-related deaths. Neoplastic cell dissemination may occur either via blood vessels or via the lymphatic system (Paduch, 2016). VEGFC is one of the key factors promoting malignant cell spread, as demonstrated both in mouse tumor models (Mandriota et al, 2001; Skobe et al, 2001; Ma et al, 2018) and during human cancer progression (Thiele & Sleeman, 2006; Rinderknecht & Detmar, 2008; Chen et al, 2012; Jiang et al, 2014). Similarly, it was shown that BACH1 promotes the metastasis of breast cancer through different molecular mechanisms (Liang et al, 2012; Lee et al, 2013; Lee et al, 2014) and its stabilization in lung adenocarcinoma is associated with increased metastatic dissemination and poor survival (Lignitto et al, 2019; Wiel et al, 2019). To substantiate the pathophysiological relevance of the interaction between BACH1 and VEGFC, we carried out an in silico analysis of publicly available gene-expression data generated by The Cancer Genome Atlas Research Network (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga). We analyzed RNA sequencing–derived data regarding aberrant gene expression in specimens taken from melanoma and lung adenocarcinoma (LUAD) human cancer patients. Melanoma is one of the commonest forms of skin cancer, whereas LUAD is at present the most common lung cancer subtype among nonsmokers. In both cancers, although the early-stage is curable by surgical resection, lymphatic metastasis results in poor prognosis. Interestingly, we found that BACH1 expression positively and significantly correlates with the expression of VEGFC in human melanoma and LUAD cancer progression (Fig 8). Specifically, BACH1 and VEGFC expression are significantly higher in samples from melanoma patients clinically diagnosed with lymph node metastatic spread, as compared with those with primary tumors (Fig 8A). Similarly, the expression of BACH1 and VEGFC were significantly augmented in specimens derived from clinical stage III LUAD patients in comparison with stage I and II (Fig 8B). According to the tumor node metastasis taxonomy classification, stage I refers to the early, nonmetastatic stage, whereas stages II and III usually indicate the intermediate, regional lymphatic metastatic stages, of which stage III has a higher lymphatic metastasis degree than stage II. This correlation may indicate the potential ability of BACH1 and VEGFC to promote cellular migration and cancer invasion. Hence, BACH1 and VEGFC may serve as candidate diagnostic biomarkers in cancer patients.

(A) Correlation of BACH1 and VEGFC expression in specimens from melanoma patients clinically diagnosed with a primary tumor (n = 84) in comparison with those with metastatic stage melanoma (n = 356), as deduced from The Cancer Genome Atlas Research Network RNA sequencing (RNA-Seq) data. Mean ± SEM; *P ≤ 0.0008. (B) Correlation of BACH1 and VEGFC expression in specimens from lung adenocarcinoma (LUAD) clinically diagnosed with primary tumor stages I (n = 292), II (n = 133), and III (n = 95), as analyzed from The Cancer Genome Atlas Research Network RNA-Seq. data. Mean ± SEM; *P < 0.02. (A, B) Wilcoxon rank sum test in panel (A) and Kruskal–Wallis test in panel (B).
Discussion
VEGFC is a potent regulator of the growth and maintenance of blood and lymphatic vessels during embryonic development, tumor expansion, and metastasis. The results presented here demonstrate that members of the BACH family regulate VEGFC expression, thereby promoting angiogenesis and lymphangiogenesis during zebrafish development and in ovarian and lung mouse tumor models.
BACH1 and BACH2 are important for the homeostasis of heme, an essential molecule for many biological functions (Warnatz et al, 2011; Igarashi & Watanabe-Matsui, 2014). Imbalanced levels of heme can cause oxidative stress when it reacts with molecular oxygen, which in turn disrupts various cellular signaling pathways. BACH1 was initially discovered as a physiological repressor of heme oxygenase-1, a rate-limiting enzyme in heme catabolism stimulated by nuclear factor erythroid 2-related factor 2 (NRF2) (Warnatz et al, 2011). Recent emerging evidence, however, indicate the widespread functions of BACH1 in diverse physiological and pathological settings, including hematopoiesis, inflammation, cardiovascular disease, aging, metabolic disorders, and neurodegenerative diseases. BACH2 was established as a crucial factor for B- and T-cell memory differentiation. Nevertheless, the potential roles of these transcription factors during embryogenesis remain obscure (Sidwell & Kallies, 2016). Members of the BACH family possess both a BTB/POZ and a CNC-bZip functional domain, conferring them with protein–protein interaction and DNA-binding capabilities (Zhou et al, 2016) to the antioxidant response elements (Zhou et al, 2016). The BTB/POZ domain, located at the N terminus, mediates dimer formation (Igarashi & Watanabe-Matsui, 2014). Conversely, the bZip domain enables DNA binding and the formation of heterodimers with small musculoaponeurotic fibrosarcoma (sMaf) transcription factors (Davudian et al, 2016a). Three evolutionary conserved sMafs have been identified in mammals and an additional one in zebrafish, all displaying complex expression patterns during embryogenesis (Yamazaki et al, 2012). Whereas single sMaf knockout mice show no or a mild phenotype, triple-knockout embryos display severe growth retardation and liver hypoplasia, resulting in embryonic lethality at E13.5 (Yamazaki et al, 2012). Interestingly, a member of the large Maf family of transcription factors, mafba in zebrafish (Koltowska et al, 2015) and Mafb in mice (Dieterich et al, 2015), was shown to be activated by VEGFC. In zebrafish, mafba is crucial for the migration of lymphatic precursors after their initial sprouting from the cardinal vein (Koltowska et al, 2015). In LECs, MAFB promotes the expression of PROX1, KLF4, NR2F2, and SOX18, key transcription factors and markers of differentiated LECs. Furthermore, E14.5 Mafb−/− mouse embryos show impaired lymphatic patterning in the skin (Dieterich et al, 2015). Altogether, these studies suggest that the BACH and MAF transcription factor families play key roles during lymphangiogenesis. Uncovering the complexity within these molecular networks may be exploited for the understanding of EC differentiation and vascular development.
Four bach genes (bach1a, bach1b, bach2a, and bach2b) with homology to mammalian BACH are present in the zebrafish genome. Phylogenetic analysis indicates that bach1a, bach1b and bach2a, bach2b belong to two distinct groups, bach1 and bach2, respectively, with bach2 possibly diverging earlier than bach1 (Luo et al, 2016). It was shown that, during embryogenesis, the two bach1 genes inhibit heme oxygenase 1a (hmox1a) induction in zebrafish (Fuse et al, 2015). In a recent study, overexpression of bach1b was found to suppress developmental angiogenesis by inhibiting Wnt/β-catenin signaling (Jiang et al, 2017). Herein, we show that the bach2a gene controls vegfc expression, directing blood and lymphatic vascular development in zebrafish. Yet, the lack of full overlapping expression between bach2a and vegfc supports additional tissue-specific functions for each of these factors.
Accumulating data establish BACH1 as a critical facilitator of tumorigenesis and metastasis in breast (Lee et al, 2013), colon (Davudian et al, 2016b), prostate (Shajari et al, 2018) ovarian (Han et al, 2019), and lung (Lignitto et al, 2019; Wiel et al, 2019) cancer. Elevated levels of BACH1 expression have been linked to a higher risk of breast cancer recurrence in patients (Liang et al, 2012), whereas association with metastatic spread and poorer prognosis has recently been suggested in the case of human ovarian cancer (Han et al, 2019) and lung adenocarcinoma (Lignitto et al, 2019; Wiel et al, 2019). Ectopic expression of BACH1 in breast cancer cells promotes malignancy and metastasis, whereas its knockdown suppresses these processes. BACH1 has been placed downstream of the Raf kinase inhibitory protein, a tumor suppressor gene shown to inhibit invasion and bone metastasis in a breast cancer xenograft mouse model (Yun et al, 2011; Lee et al, 2013). Inactivation of Raf kinase inhibitory protein during tumor expansion results in higher expression of BACH1 and its target genes C-X-C chemokine receptor type 4 (CXCR-4) and matrix metalloproteinase1 (MMP1), established drivers of tumor progression and metastasis (Foley & Kuliopulos, 2014; Mishan et al, 2016). Furthermore, ablation of BACH1 in human colon carcinoma (Davudian et al, 2016b) or in prostate cancer cells (Shajari et al, 2018) prevents cell growth, migration, and invasion in vitro, decreasing the expression of its main metastasis-related genes, MMP1, let-7a, and CXCR4. Interestingly, cxcr4 is expressed in the somites and the endothelium of zebrafish embryos (Chong et al, 2001), where we detect the expression of both bach2a and bach2b transcripts. cxcr4 was shown to be crucial for lateral aortae formation (Siekmann et al, 2009) and trunk lymphatic vascular network assembly (Cha et al, 2012) and a key modulator of vascular progenitor cells (Sainz & Sata, 2007). In addition to its contribution to development, CXCR4 was shown to play a role in carcinogenesis, tissue repair and other pathological circumstances (Kawaguchi et al, 2019). VEGFC can, thus, be added to a growing list of tumor and metastatic proteins, including CXCR4 and MMP1, all of which are transcriptionally regulated by BACH1. Including VEGFC as a target of BACH1 allows novel perspectives of the role of BACH1 in vascular development during embryogenesis and pathological conditions.
During the past years, a substantial amount of in vitro, animal, and small-size human studies established BACH proteins as a hub of critical transcriptional networks that govern key processes during normal physiology and disease states. Surprisingly, however, Bach single-(Sun et al, 2002; Muto et al, 2004) or double- (Ebina-Shibuya et al, 2016) knockout mice exhibit no phenotype at birth. This may result from the existence of a yet unknown compensatory mechanism similar to the one described here in zebrafish. Further understanding the mechanisms underlying BACH’s mode of action during embryogenesis, adult life, and tumorigenesis would pave the way to resolving this ambiguity.
Altogether, our results highlight BACH as a novel regulator of angiogenesis and lymphangiogenesis during embryonic development as well as tumor progression. We show that BACH controls the expression of VEGFC, an established pro-lymphangiogenic and angiogenic growth factor. The expanded recognition of BACH1 activity, together with its inhibitory effect on NRF2, an important detoxifying and antioxidant factor, marks it as a potential therapeutic target. Hence, various direct or indirect promising BACH1 modulators have been developed (Arbiser, 2011; Banerjee et al, 2013; Cuadrado et al, 2019). Accordingly, recent reports demonstrated that targeting of BACH1 and mitochondrial metabolism may serve as an effective therapy for triple negative breast cancer (Lee et al, 2019). In two new studies on lung cancer, it was shown that therapeutic intervention that either destabilizes BACH1 (Lignitto et al, 2019) or disrupts its ability to induce glycolysis (Wiel et al, 2019) have the potential to inhibit BACH1 pro-metastatic activity. Targeting the BACH1/VEGFC signaling axis with these inhibitors may potentially be significant therapeutically for various blood and lymphatic vessels pathologies as well.
Materials and Methods
Bioinformatics analysis
Transcription factor–binding site analysis was performed using the Genomatix Genome Analyzer (Genomatix Software GmbH) MatInspector program (Cartharius et al, 2005). Promoter regions of the human, mouse, and zebrafish Vegfc genes were extracted from the University of California Santa Cruz genome browser (Kent et al, 2002) as follows: Human-GRC37/hg19 chr4: 177,713,306-177,716,211; Mouse-GRCm38/mm10 chr8:54,075,150-54,077,946; and Zebrafish Zv9/danRer7 chr1:39,270,126-39,273,225. The zebrafish bach sequences used are bach1a: NM_001040313.1; bach1b: NM_001020663.1; bach2a: XM_680223.9; and bach2b: XM_677841.6.
Zebrafish lines and husbandry
The EK and Tg(fli1:EGFP)y1 (Lawson & Weinstein, 2002) zebrafish lines were maintained under standard conditions (Isogai et al, 2003). All experiments were carried out according to the guidelines of the Weizmann Institute Animal Care and Use Committee.
FACS and RT-PCR analysis
Tg(fli1:EGFP)y1 embryos at 21–24 hpf and 3 dpf were collected and dissociated as previously described (Nicenboim et al, 2015). FACS of single-cell suspensions was performed at 4°C using FACS Aria flow cytometer (Becton Dickinson). Total RNA was isolated from equal numbers of GFP+ and GFP− cells by PerfectPure RNA Cultured Cell kit (5 PRIME), according to the manufacturer’s instructions. RNA (100 ng) from each sample was subjected to first-strand cDNA synthesis with Superscript III reverse transcriptase (Invitrogen) and random hexamers. All PCR conditions were optimized to produce a single product of the correct base pair size in the linear range of the reaction using the following set of primers: bach1a: 5′-TGTAAGACGGCGGAGTAAGA and 5′-CTTCAGCTGGTTGTGGTCT, bach1b: 5′-CTTCAGTGCTCGTGTGTCCA and 5′-TGTAGGCGAACTCCAGCAAG, bach2a: 5′-GACAGAACACGAGCCACTCA and 5′-ACAGCGCATGACATCTTGGA, bach2b: 5′-TGCATCCTGAACCTTGAGTGT and 5′-CTGCACATCTCGACACACCT, fli1: 5′-CCGAGGTCCTGCTCTCACAT and 5′-GGGACTGGTCAGCGTGAGAT, bactin: 5′-CGAGCAGGAGATGGGAACC and 5′-CAACGGAAACGCTCATTGC.
Construction of the pCS2vegfcCDS plasmid
The full-length coding region of zebrafish vegfc was amplified from cDNAs derived from 24 hpf embryos using a forward (5′-ATGCACTTATTTGGATTTTCT) and a reverse (5′-TTAGTCCAGTCTTCCCCAG) primers, and sub-cloned into the pCRII-TOPO cloning vector (Invitrogen). After nucleotide sequence verification, a Gateway-compatible (Invitrogen) middle entry clone was generated using Gateway BP clonase (Invitrogen)–mediated recombination. A pCS2vegfcCDS plasmid was produced using Gateway LR clonase (Invitrogen). Capped mRNA was transcribed from a Not1-linearized template using the mMESSAGE mMACHINE Kit (Thermo Fisher Scientific).
Microinjection of zebrafish embryos
Morpholino antisense oligonucleotides (Gene Tools) were designed with sequences complementary to zebrafish bach2a and bach2b cDNA in a location downstream to the initiating start codon. The morpholino sequences were as follows: bach2a: (GenBank accession number XM_680223.9), 5′-TGTCAGGCTTCTCCTCCATAGACAT-3′ and bach2b: (GenBank accession number XM_677841.6), 5′-CTTCAGACTTCTCATCCACGGACAT-3′. The morpholino targeting the zebrafish vegfc was previously described (Yaniv et al, 2006). As a control, the Gene Tools standard control MO (5′-CCTCTTACCTCAGTTACAATTTATA-3′) was applied. Specific MOs were injected into one-cell stage embryos at concentrations from 3.75 up to 10 ng and control MO at 10 ng per embryo. For rescue experiments, pCS2vegfcCDS mRNA (800 pg per embryo) was simultaneously injected with the morpholino (3.75 ng per embryo). For quantification of the phenotypes, embryos from each group were randomly selected. Quantification was at least three independent experiments.
Design and synthesis of gRNA
The design of the bach2a and bach2b CRISPR guide was performed with CHOPCHOP (Montague et al, 2014) (chopchop.rc.fas.harvard.edu). Potential off-target sequences were checked using the MIT CRISPR Design site (crispr.mit.edu) (Hsu et al, 2013). Oligonucleotides synthesized for the guide sequence 5′-GGACGTCCTGTGTGACGTGA and 5′-CTGTGCCGAATTCCTGCGCA for bach2a and bach2b, respectively, were cloned into the BsmBI site of the pT7-gRNA plasmid (a gift from Wenbiao Chen) (plasmid # 46759; Addgene) (Jao et al, 2013). Alt-R S.p. Cas9 Nuclease 3NLS (250 ng/μl; Integrated DNA Technologies) and gRNA (125 ng/μl) in 5 μl total volume were co-injected into one-cell–stage Tg(fli1:EGFP)y1 embryos. To detect mutagenic events, DNA was extracted from 24-hpf embryos, amplified using a set of primers 5′-AGCAAGGAATGTCTATGGAGGA and 5′-ATGAGTGGCTCGTGTTCTGTC (235 bp) for bach2a and 5′-CGCTCCATTGTTACAGTTTGC and 5′-GCCGTCCTCTTCACTGCGC (157 bp) for bach2b. PCR products were separated on 3.5% MetaPhor Agarose gel (Lonza). When both genotyping and phenotypic analyses of single zebrafish embryos from heterozygous bach2a intercross was needed, the larval tail (6 dpf) was used for genotyping, whereas RNA was extracted from the anterior part using NucleoSpin RNA Plus XS RNA purification kit (Macherey-Nagel).
Whole-mount in situ hybridization
Embryos were dechorionated and fixed overnight in 4% PFA at 4°C, at the appropriate time points. In situ hybridization was performed as previously described (Yaniv et al, 2006) using single-stranded digoxygenin-dUTP–labeled RNA probes transcribed by T7 RNA polymerase (Roche). The PCR-generated probes were amplified with the following set of primers: bach2a: 5′-AAGAGTGAGCTAGAGGGCA and 5′-CGTTCTCTTGTTCGGGATCTTG; bach2b: 5′-CTGCGCAGTGAAGAGGACGG and 5′-GCTCCACCTCTTGCTTGCAC; vegfc: 5′-CATCAGCACTTCATACATCAGC and 5′-GTCCAGTCTTCCCCAGTATG; lyve1: 5′-GGTTTGGTTGGGTTGAGGAGC and 5′-TTAGGAAGAGTCAGAGTCTTGTTC; flt4: 5′-CTCGAGAATGACATGTGCTGG and 5′-CAGCCAGCGAGCACAAAGC. After performance of a color reaction with alkaline phosphatase substrates (Roche), embryos were fixed in 4% PFA and washed in PBS supplemented with 0.1% Tween-20 (PBST). For tissue clarification, embryos were mounted in glycerol and imaged using Leica M165 FC stereomicroscope.
Cell culture
Human ovarian clear cell carcinoma ES2 cell line (American Type Culture Collection) was cultured in DMEM supplemented with 10% FBS. Mouse D122 Lewis lung carcinoma cell line (Eisenbach et al, 1984), kindly provided by Prof. Lea Eisenbach, (Weizmann Institute of Science, Israel), was grown in DMEM containing 10% FBS, 1 mM sodium pyruvate, and 1% nonessential amino acids. All cell lines were routinely tested for mycoplasma contamination using the EZ-PCR Mycoplasma Test Kit (Biological Industries).
Establishment of stable BACH1-overexpressing cell pools
Full-length coding region of the human BACH1 (BACH1, GenBank accession number NM_206866.3) and mouse Bach1 (Bach1, GenBank accession number BC057894.1) with an N-terminal HA tag were reverse transcribed with SuperScript III Reverse Transcriptase (Invitrogen) and PCR-amplified using Phusion high-fidelity DNA polymerase (New England Biolabs) together with the following set of primers: BACH1: 5′-ATGTCTCTGAGTGAGAACTCGG and 5′-TTACTCATCAGTAGTACATTTATC; Bach1: 5′-ATGTCTGTGAGTGAGAGTGCG and 5′-TTACTCGTCAGTAGTGCACTTG. The fragments were ligated into pCRII-TOPO and their sequence fidelity was confirmed by sequencing. Inserts were restricted and ligated into pEIRES expression vector containing the human EF-1a promoter (Hobbs et al, 1998) to produce the pIRESBACH1 and pIRESBach1 constructs. ES2 and D122 cells were transfected with pIRESBACH1 and pIRESBach1 expression vectors, respectively, using Lipofectamine 2000 reagent (Invitrogen) and selected with 2.5 μg/ml puromycin (Sigma-Aldrich) as was previously described (Cohen et al, 2009). An average of 50 individual puromycin-resistant colonies were collected together, and BACH1 overexpression was confirmed by immunoblotting. ES2 and D122 cells that were stably transfected with the pEIRES empty vector were used as controls.
ChIP assay
The ChIP experiments were performed using the ChIP-IT-Express kit (Active Motif Cat. no. 53009). Cells were fixed and cross-linked using 1% formaldehyde at room temperature for 10 min. Fixation was stopped by adding glycine. DNA extracted from nuclear fraction was subjected to enzymatic shearing for 35 min at 37°C to obtain mononucleosomes. The resulting chromatin preparation was immunoprecipitated with magnetic protein G–coupled beads and 10 mg of either anti HA-tag (HA.11; Covance), Bach1 (sc-14700; Santa Cruz), or nonspecific-IgG antibody. DNA–protein crosslinking was reversed at 65°C (4 h), treated with Proteinase K (2 h, 42°C). The recovered DNA was then subjected to PCR using the following set of specific primers: BACH proximal site: 5′-GAGGGAGAGTGAGAGGGG and 5′-CGCAGGATCCTCCAGAGC; BACH distal site: 5′-CCGAGTCTGATGGGATGGAA and 5′-GCCTTTGTTGATACAGCCTTGG.
Transient transfection and luciferase assay
The 5′ regulatory region of human VEGFC gene (NG_034216.1) encompassing 2,274 nucleotides (−1 to −2,274 in relation to ATG, Fig 1) was synthesized by GenScript. This fragment was then used as a template for deletion of either the proximal (nt −623 to −603, 5′-GGCGCGTCAGTCATGCCCTGC) or distal (nt −2074 to −2054, 5′-GTAGTGTGAGTCAA CACTGTG) BACH-binding site individually or in combination. Subsequently, these synthesized fragments were sub-cloned into the pGL4.10[luc2] promoter-less vector (Promega) between KpnI and XhoI restriction sites to generate the pVEGFCwt-Luc (wild-type VEGFC promoter region), pVEGFCΔPro-Luc (VEGFC promoter region deleted of the proximal BACH-binding site), pVEGFCΔDis-Luc (VEGFC promoter region deleted of the distal BACH-binding site), and pVEGFCΔProDis-Luc (VEGFC promoter region deleted of both proximal and distal BACH-binding sites). All constructs were verified by DNA sequence analysis. For the luciferase reporter gene assay, ES2 cells plated in 24-well plates (40,000 cells per well) were co-transfected with the indicated pVEGFC-Luc construct in combination with the pRL-TK-Renilla (pRL-TK vector; Promega) luciferase internal control vector using Lipofectamine 2000 (Thermo Fisher Scientific), according to the manufacturer’s instructions. After 48 h, the cells were lysed and luciferase activity was measured using the Dual-Luciferase Assay System (Promega). All luciferase data were corrected for transfection efficiency based on the Renilla internal control following by subtraction of the pGL4.10[luc2] activity background. The data were calculated as means ± standard error of three independent experiments, each performed in nine replications. Luciferase activities for each transfection are plotted as average fold-change in relation to the pVEGFCwt-Luc.
Western blot analysis
Protein extraction was performed in RIPA buffer (20 mM Tris, pH 7.4, 137 mM NaCl, 10% glycerol, 0.5% [wt/vol] sodium deoxycholate, 0.1% [wt/vol] SDS, 1% Triton X-100, and 2 mM EDTA) supplemented with 1 mM PMSF and protease inhibitor cocktail (Sigma-Aldrich). Lysates (20 μg/lane) were electrophoresed in SDS–PAGE under reducing conditions and transferred to a nitrocellulose (Whatman). Membranes were probed with anti HA-tag monoclonal antibody (HA.11, 1:100; Covance), whereas β-tubulin (H-235, 1:500; Santa Cruz) was used as a loading control. Appropriate HRP-conjugated antimouse or antirabbit secondary antibodies (1:10,000; Jackson ImmunoResearch) were used, respectively.
Cell invasion assay
The invasion potential of the cells was examined in vitro using BioCoat Matrigel Invasion Chamber (Corning). A total of 80 × 103 cells suspended in serum-free medium were seeded in the upper chamber. To initiate cell invasion, medium supplemented with 10% FBS was added as a chemoattractant in the low chamber. The cells were incubated at 37°C for 20 h and invaded cells on the inferior surface of the inserts were fixed, washed, and stained with crystal violet (Sigma-Aldrich). Each assay was repeated two times in duplicates. For analysis, the entire Matrigel surface area was imaged (Olympus SZX16 stereomicroscope, 2.5× magnification) and quantified using the ImageJ software. To ensure that the difference in invasion rate is not due to differential cell growth rate, total cell number validation was carried on a parallel plate. Cells were stained with crystal violet, lysed with 1% SDS, 0.1N NaOH solution followed by direct dye intensity measurement.
Tumor initiation, histologic preparation, immunohistochemistry, and morphometric analysis
Subcutaneous tumors were generated by injecting 2 × 106 single-cell suspensions of cells in 100 μl PBS into a shaved lower right flank of 7-wk-old mice. D122 Control or Bach1-overexpressing cells were injected to male immunocompetent syngeneic C57BL/6 (Harlan Laboratories), whereas ES2 cells were injected to female immunodeficient CD1 nude (Harlan Laboratories) mice. Orthotopic metastatic tumor growth was initiated by injecting 1 × 106 ES2 cells intraperitoneally at a remote site in the abdomen of 7-wk-old female CD1 nude mice (Harlan Laboratories). In all experiments, animals were randomly assigned to the control and overexpressing cells injected groups. Excised mouse tumors were rinsed in ice-cold PBS and then gradually fixed at 4°C in 2.5% and 1% PFA for 24 and 48 h, respectively. Paraffin-embedded tissue was sectioned serially at 4 μm thickness. The first slide was stained with hematoxylin and eosin, whereas other representative slides underwent immunohistochemical staining using the following antibodies: anti CD34 (1:100; Cedarlane Laboratories), LYVE1 (1:100; Fitzgerald Industries), cytokeratin 7 (ab9021, 1:200; Abcam), and VEGFC (H-190 1:200; Santa Cruz). Morphometric blood and lymphatic vessel coverage analysis were performed on CD-34 and LYVE1 stained tumor sections, respectively. Images of CD-34–stained sections were captured with a fluorescence microscope (NI-U; Nikon), equipped with Plan Fluor objectives connected to CCD camera (DS-Ri1; Nikon). Digital images were collected and assembled using Adobe Photoshop (Adobe Systems). LYVE1 immunohistochemical-stained sections were scanned using the Panoramic Viewer. The density of vessels was evaluated using Image Pro Plus software (Media Cybernetics). For all analysis, investigators were blinded and unaware of group allocation. All animal experiments described in this study were performed according to the guidelines of the Weizmann Institute Animal Care and Use Committee. For immunostaining of ES2 cells, fixation was carried out in 4% paraformaldehyde for 10 min. Fixed cells were then blocked for unspecific staining in 0.3% Triton X-100, 10% horse serum for 90 min at RT, incubated with antibodies directed either against HA-tag (1:200; Sigma-Aldrich) or VEGFC (H-190 1:200; Santa Cruz) antibody for 2 h at room temperature, and visualized by incubating the cells with goat anti-rabbit Cy3 (1:10,000; Jackson ImmunoResearch) secondary antibody. Nuclei were counterstained with DAPI (Invitrogen).
Quantitative real-time-PCR
Total RNA was isolated from cultured cells or tumor specimens by PerfectPure RNA Cultured Cell or Tissue kit (5 PRIME), respectively, according to the manufacturer’s instructions. RNA (1 μg) from each sample was subjected to first-strand cDNA synthesis with the High-Capacity cDNA Reverse Transcription kit (Applied Biosystems) and random hexamers. Quantitative real-time PCR was conducted with LightCycler-FastStart DNA Master SYBR Green I kit (Roche) using a LightCycler 480 real-time PCR System (Roche). The relative expression level of each target gene was determined using GAPDH and beta-2-microglobulin (B2M) as reference genes. Primers used were as follows: human BACH1: 5′-TCTTCCAGAAGAGGTGACAGT and 5′-ACTCCACACATTTGCACACT; VEGFA: 5′-ATGCGGATCAAACCTCACC and 5′-T CTTTCTTTGGTCTGCATTCAC; VEGFB: 5′-CCACCAGAGGAAAGTGGTGTC and 5′-ACAGCGCTGCACAGTCAC; VEGFC: 5′-GCCACGGCTTATGCAAGCAAAGAT and 5′-AGTTGAGGTTGGCCTGTTCTCTGT; MMP1: 5′-CTGGCCACAACTGCCAAATG and 5′-CTGTCCCTGAACAGCCCAGTACTTA; GAPDH: 5′-AGGGCTGCTT TTAACTCTGGT and 5′-CCCCACTTGATTTTGGAGGGA; B2M: 5′-TTCTGGCCT GGAGGCTATC and 5′-TCAGGAAATTTGACTTTCCATTC; Mouse Bach1: 5′-TGACAGCGAGTCCTGTTCTG and 5′-TTATCCGTTGGGCATTGAA; Vegfa: 5′-TCT TCAAGCCATCCTGTGTG and 5′-GAGGTTTGATCCGCATAATCTG; Vegfb: 5′-ACGATGGCCTGGAATGTGTG and 5′-TGGTCTGCATTCACATTGGC; Vegfc: 5′-GTAAAAACAAACTTTTCCCTAATTC and 5′-TTTAAGGAAGCACTTCTGTGTGT; Gapdh: 5′-GACGGCCGCATCTTCTTGTG and 5′-CTTCCCATTCTCGGCCTTGACTGT; B2m: 5′-CCCGCCTCACATTGAAATCC and 5′-GCGTATGTATCAGTCTCAGTGG; Zebrafish bach2b; 5′-CAGCATGCCAGAGGAGGT and 5′-AGTGATTGCTCTCCGACGC; and bactin 5′-TGACAGGATGCAGAAGGAGA and 5′-GCCTCCGA TCCAGACAGAGT.
Clearing and immunofluorescence staining
Excised tumors were fixed with 4% PFA for 1 wk at 4°C, washed three times with PBS, permeabilized with (0.2% Triton X-100 in PBS) for 4 h, and immersed overnight in blocking solution (PBS containing 0.05% Triton X-100 and 10% normal goat serum). For immunofluorescent staining, tumors were incubated with primary mouse anti-LYVE1 antibody (Fitzgerald Industries) diluted (1:250) in an antibody cocktail (50% blocking solution, 0.05% Triton X-100 in PBSX1) for 1 wk at 4°C, washed 24 h (1% blocking solution, 0.05% Triton X-100 in PBS), and probed with an Alexa Flour 594–conjugated goat antirabbit secondary antibody (A11037, 1:250; Molecular Probes—Life Technologies) diluted in antibody cocktail for an additional week at 4°C. After 24-h wash, the tumors were subjected to a Whole Organ Blood and Lymphatic Vessels Imaging (WOBLI) clearing procedure (Hama et al, 2011; Chung et al, 2013; Oren et al, 2018). Briefly, the tumors were re-fixed with 4% PFA for 24–72 h at 4°C, transferred to hydrogel solution (4% Acrylamide, 0.025% Bis-acrylamide, 0.25% Va-044 and 4% PFA in PBS) for 1 wk, and passively cleared (200 mM Boric acid and 4% SDS) for 2 wk at 37°C. Subsequently, the tumors were placed in Scale solution (4M urea, 10% glycerol and 0.1% Triton X-100) for 48 h. A sequence of 3D images and movies were acquired using Zeiss LSM710 confocal microscope (Carl Zeiss).
Microscopy
Images were acquired on Zeiss LSM 780 upright confocal microscope (Carl Zeiss) with a W-Plan Apochromat ×20 objective, NA 1.0. Fluorescent proteins were excited with single-photon laser (488 nm). Alternatively, Leica TCS SP8 microscope, equipped with environmental control, two internal Hybrid (HyD) detectors, and Acusto Optical Tunable Filter (Leica microsystems CMS GmbH) was used, and excitation was performed using 488-nm Ar laser and emission was collected using the internal HyD detector at 510–625 nm, with a gain of 100.
Statistical analysis
Statistical analyses were performed with analytic computerized software (Statistix 8 Student Edition, Analytical Software). Comparisons between treatment groups were performed with either ANOVA or Kruskal–Wallis nonparametric ANOVA (alternatively, t test or Wilcoxon rank sum test were used if only two groups were compared). ANOVA test was used to analyze normally distributed data (evaluated by Shapiro–Wilk test) that had equal variances between groups (evaluated by Bartlett’s test), whereas Kruskal–Wallis nonparametric ANOVA was used to analyze data that were not normally distributed and/or had unequal variance between groups; when relevant, Tukey HSD test or mean ranks test, respectively, were used for all pairwise comparisons. Differences were considered significant at P < 0.05. Unless otherwise noted, data are presented as mean ± SEM.
Acknowledgements
The authors would like to thank Irit Orr and Ron Rotkopf for assistance with human cancer data analysis; Prof Eytan Domany and Dr Noa Ben-Moshe for transcription factor analysis; Ayala Sharp for FACS analysis support; Michal Shemesh for assisting with imaging requirements; Ofir Atrakchi, Yona Ely, Ron Hadas, Tal Lupo, and Gila Meir for technical assistance; Noa David for graphic design; Prof Lea Eisenbach for providing the mouse D122 Lewis lung carcinoma cell line; Gabriella Almog, Ala Glozman, and Roy Hofi for fish care; and Beni Siani and Alon Harmelin for animal care. This work was supported in part by the Israel Science Foundation grants 326/14 (to M Neeman and B Cohen), 748/2009 (to K Yaniv), and 861/2013 (to K Yaniv); European Research Council Advanced grant 232640-IMAGO (to M Neeman) and ERC Starting grant 335605 (to K Yaniv); the Thompson Foundation (to M Neeman); National Institutes of Health grant (R01 CA75334, to M Neeman); Marie Curie Actions-International Reintegration grants FP7-PEOPLE-2009-RG 256393 (to K Yaniv); the H&M Kimmel Institute for Stem Cell Research (to K Yaniv); and the Estate of Emile Mimran (SABRA program) (to K Yaniv). Michal Neeman is the incumbent of the Helen and Morris Mauerberger Chair in Biological Sciences. K Yaniv is supported by the Willner Family Center for Vascular Biology; the estate of Paul Ourieff; the Carolito Stiftung; Lois Rosen, Los Angeles, CA; Edith Frumin; the Fondazione Henry Krenter; the Wallach Hanna & Georges Lustgarten Fund; and the Polen Charitable Trust.
Author Contributions
B Cohen: conceptualization, data curation, and writing—original draft.
H Tempelhof: data curation and investigation.
T Raz: formal analysis, validation, and writing—review and editing.
R Oren: data curation, formal analysis, methodology, and writing—review and editing.
J Nicenboim: data curation, investigation, and methodology.
F Bochner: data curation, investigation, visualization, and methodology.
R Even: data curation.
A Jelinski: data curation.
R Eilam: data curation, visualization, and methodology.
S Ben-Dor: formal analysis, methodology, and writing—review and editing.
Y Addadi: investigation and visualization.
O Golani: software, formal analysis, and visualization.
S Lazar: data curation and methodology.
K Yaniv: conceptualization, supervision, funding acquisition, project administration, and writing—review and editing.
M Neeman: conceptualization, funding acquisition, project administration, and writing—review and editing.
Conflict of Interest Statement
The authors declare that they have no conflict of interest.
- Received February 2, 2020.
- Revision received February 23, 2020.
- Accepted February 24, 2020.
- © 2020 Cohen et al.
This article is available under a Creative Commons License (Attribution 4.0 International, as described at https://creativecommons.org/licenses/by/4.0/).
References




{kind=link}
{kind=link}
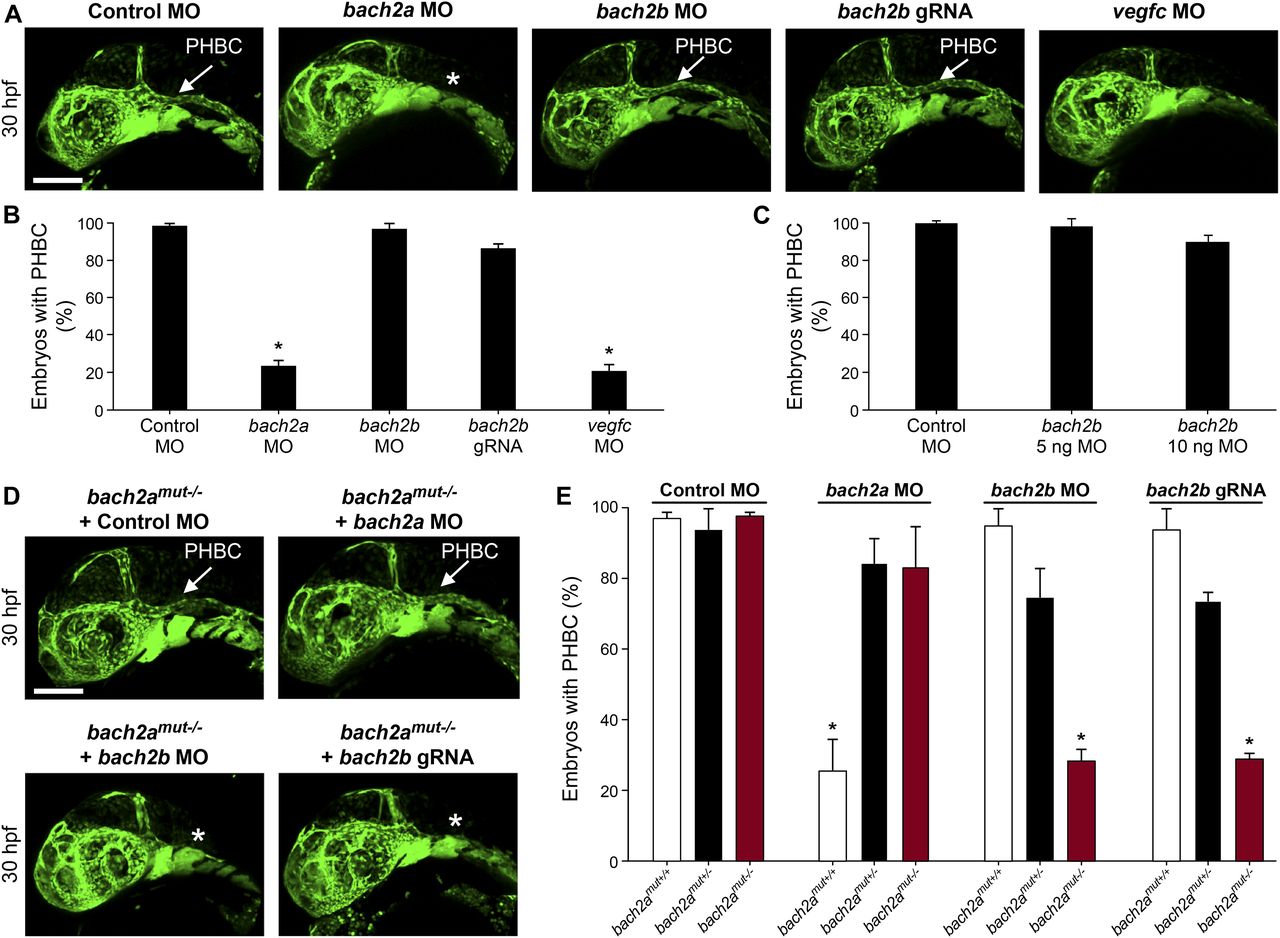{kind=link}
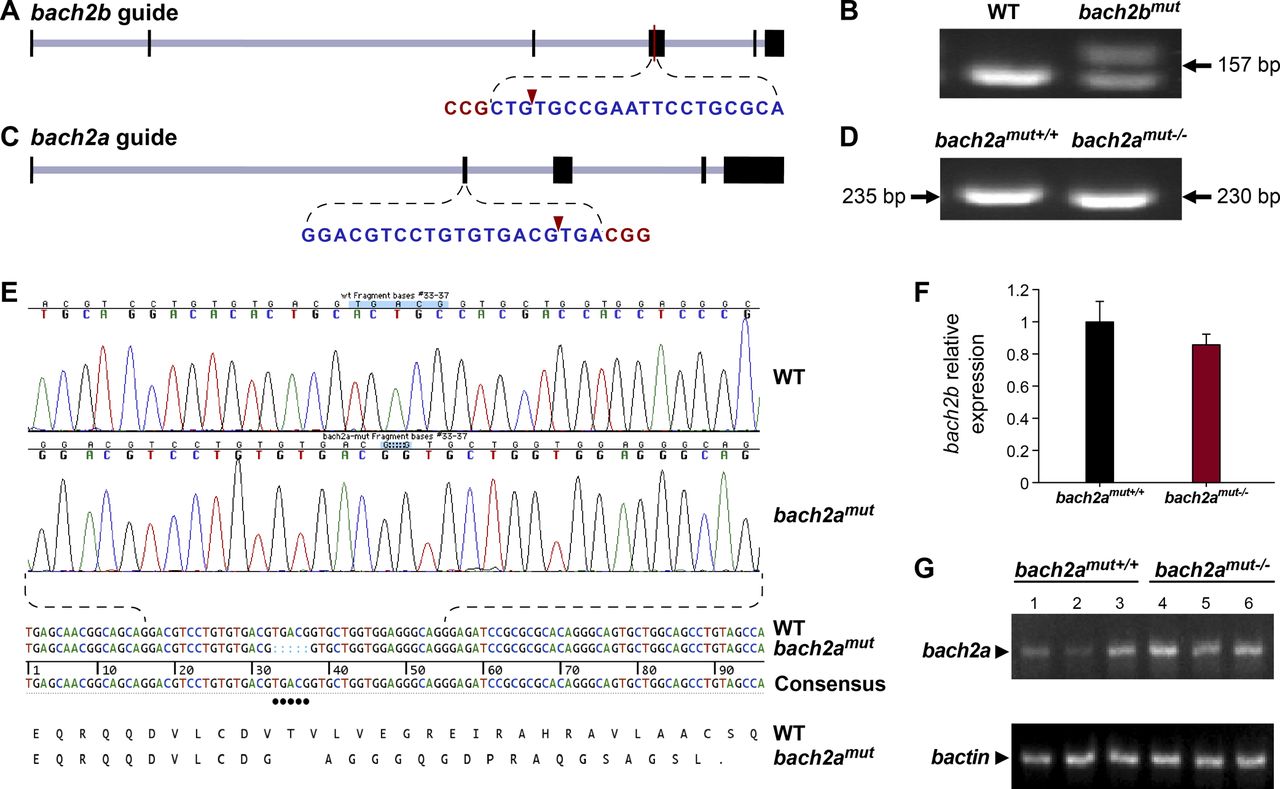{kind=link}
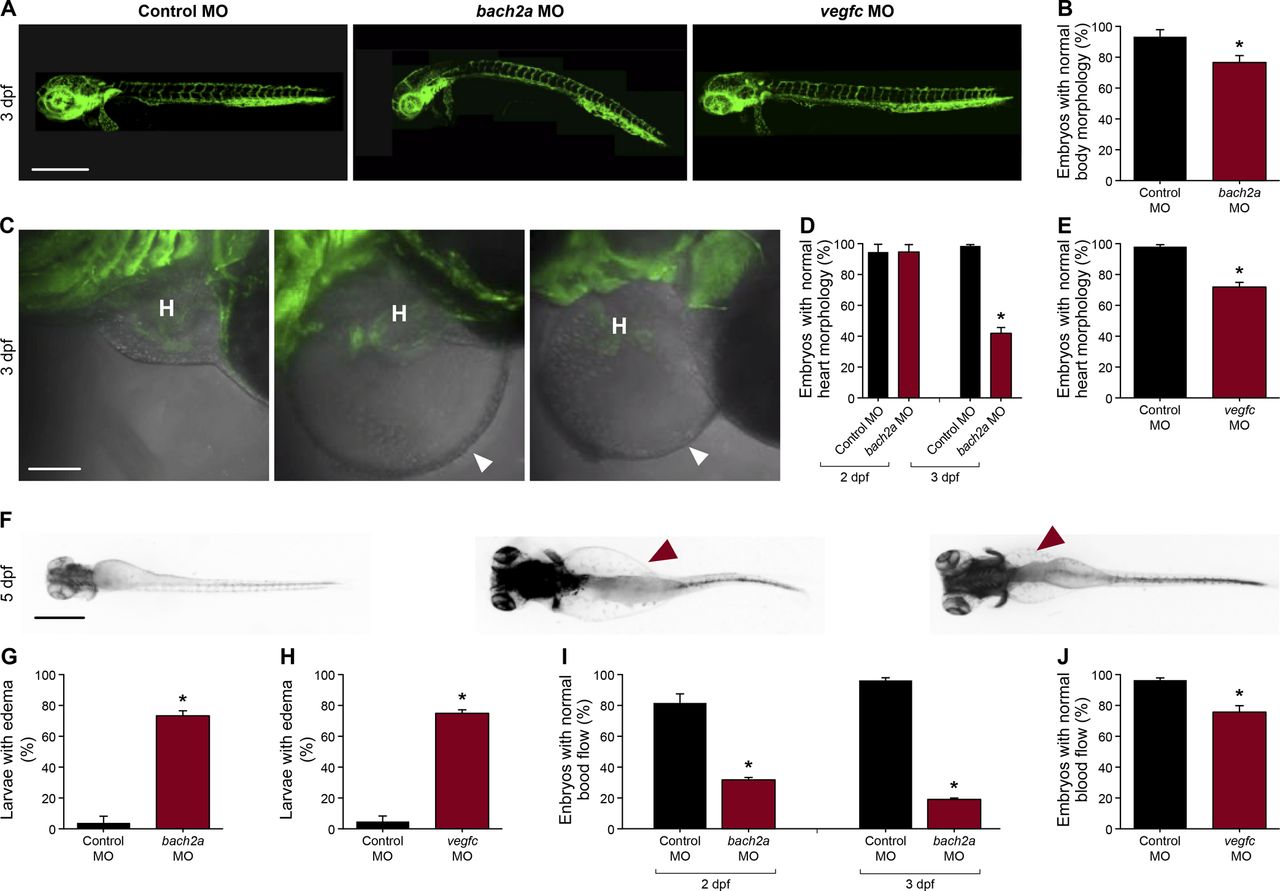{kind=link}
{kind=link}
{kind=link}
{kind=link}
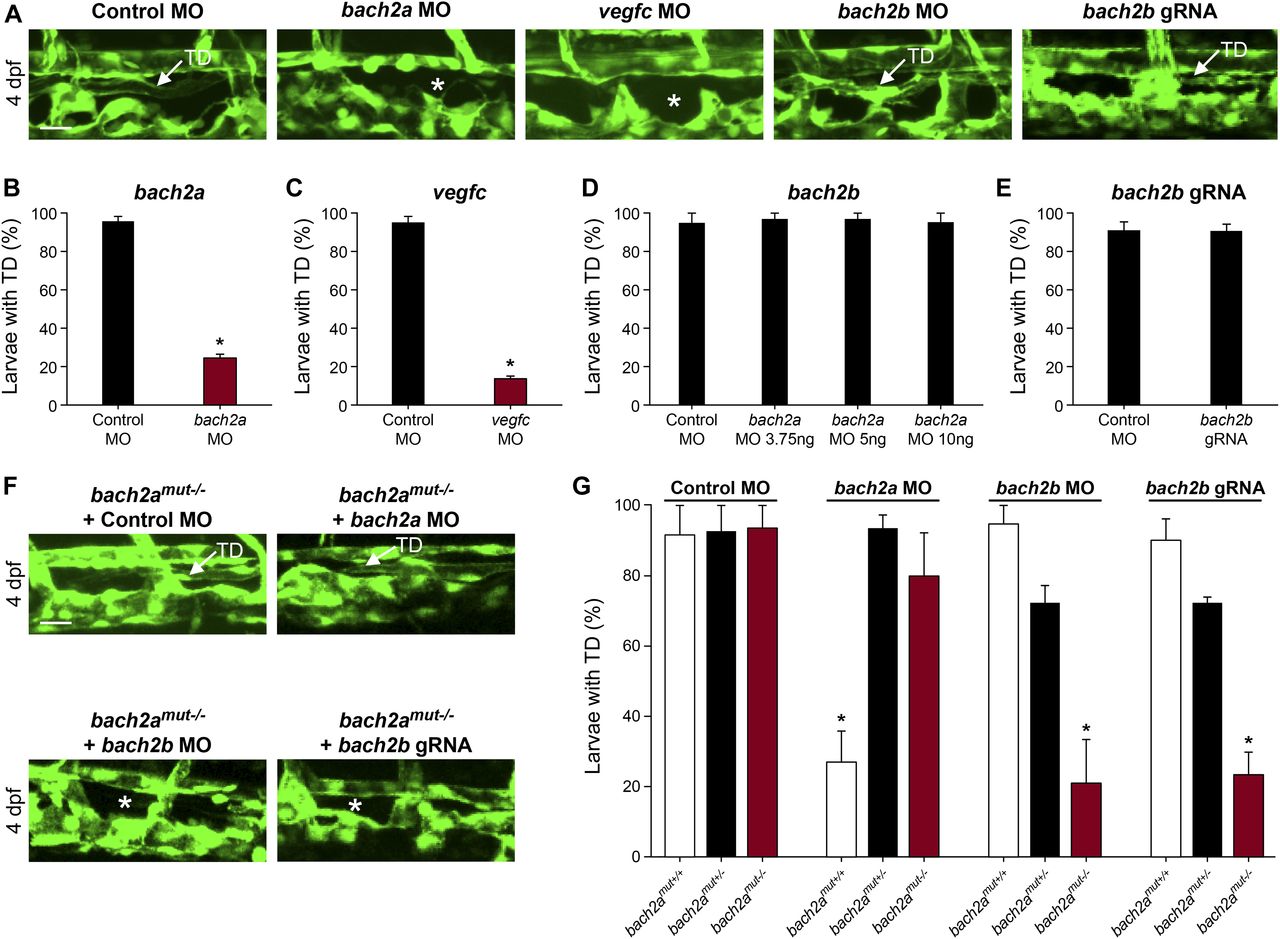{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
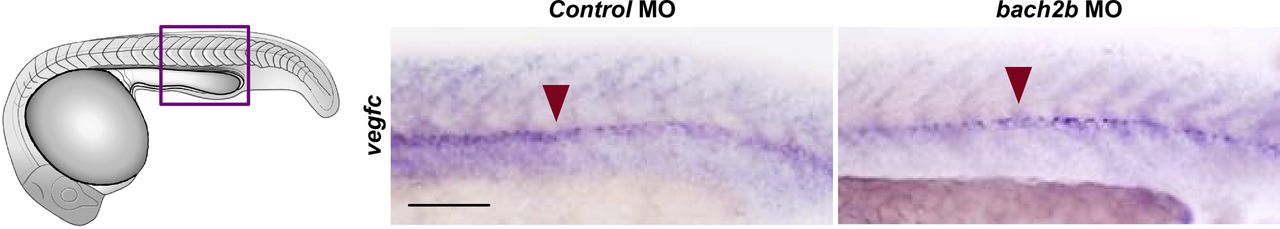{kind=link}
{kind=link}
{kind=link}
{kind=link}
