



Article Figures & Data
Figures
- Figure 1. Immunomodulatory effects of PUFAs on Drosophila inflammatory tumours.
(A) Inflammatory melanotic tumour model relies on temperature conditional activation of JAK/STAT in hopTum gain-of-function heterozygous females. (B) Melanotic tumour indicated (dashed circle). Dissected melanotic tumour from hopTum female carrying the lamellocyte marker (MSNF9-mCherry [Tokusumi et al, 2009a, 2009b]) reveals melanotic tumour is composed of lamellocyte aggregates. (C) Real-time RT-PCR was used to determine levels of inflammation markers in control and hopTum larvae raised in both standard media and media supplemented with 9-(S)-HODE (1 μM). RpL32 provided an endogenous control to normalise expression. (D, E, F) Melanotic tumour incidence was assayed in hopTum heterozygous females raised on standard medium supplemented with fatty acids. (D) SFA supplementation (coconut oil) did not increase tumour incidence. (E) Omega-6 PUFA linoleic acid (LA) supplementation increased tumour incidence. (F) Omega-3 PUFA (α-linoleic acid [ALA]) supplementation suppressed melanotic tumours even when 0.15% of LA was present. Cultures were incubated at 27°C except for omega-6 PUFA, which was conducted at both 25°C and 27°C. As hopTum is temperature-sensitive-conditional, changes in culture temperatures modify basal melanotic tumour incidence, allowing better resolution of either suppressors or enhancers of inflammation (Badenhorst et al, 2002). (G) Schematic showing biosynthetic pathways of omega-6 PUFA derived lipid mediators. The 18-carbon (C18) omega-6 PUFA LA can be converted to arachidonic acid, the precursor of eicosanoids, a family of potent bioactive lipids that include the cyclooxygenase (COX)-derived prostaglandins. (H, I) hopTum larvae were raised on media supplemented with 0.15% LA and the lipid extract was analysed by LC-MS/MS for the presence of PGF2α, PGE2, and PGD2. (H, I) Neither PGF2α (multiple reaction monitoring [MRM]: m/z: m/z 353>309, 353>247 and 353>193) nor (I) PGE2 and PGD2 (MRM: m/z 351>315, 351>271, and 351>333) were detected in Drosophila lipid extracts. Data information: in (D, E, F), 10 replicate crosses were used for each assay point. Box and whiskers plots were generated using R. * indicates values statistically significantly different from unsupplemented, P-value < 0.001 determined using t test. NS indicates not statistically significantly different.
- Figure S1. Drosophila lack prostaglandins.
(A, B) LC-MS/MS spectra for analyses to detect (A) PGF2α and (B) PGE2 and PGD2. hopTum larvae were raised on normal media (control) and media supplemented with either 0.15% LA (Omega-6) or 0.15% ALA (Omega-3). Molecular ions used were as follows: m/z 353>309, 353>193, and 353>247 for PGF2α and m/z 351>315, 351>271, and 351>333 for PGE2 and PGD2.
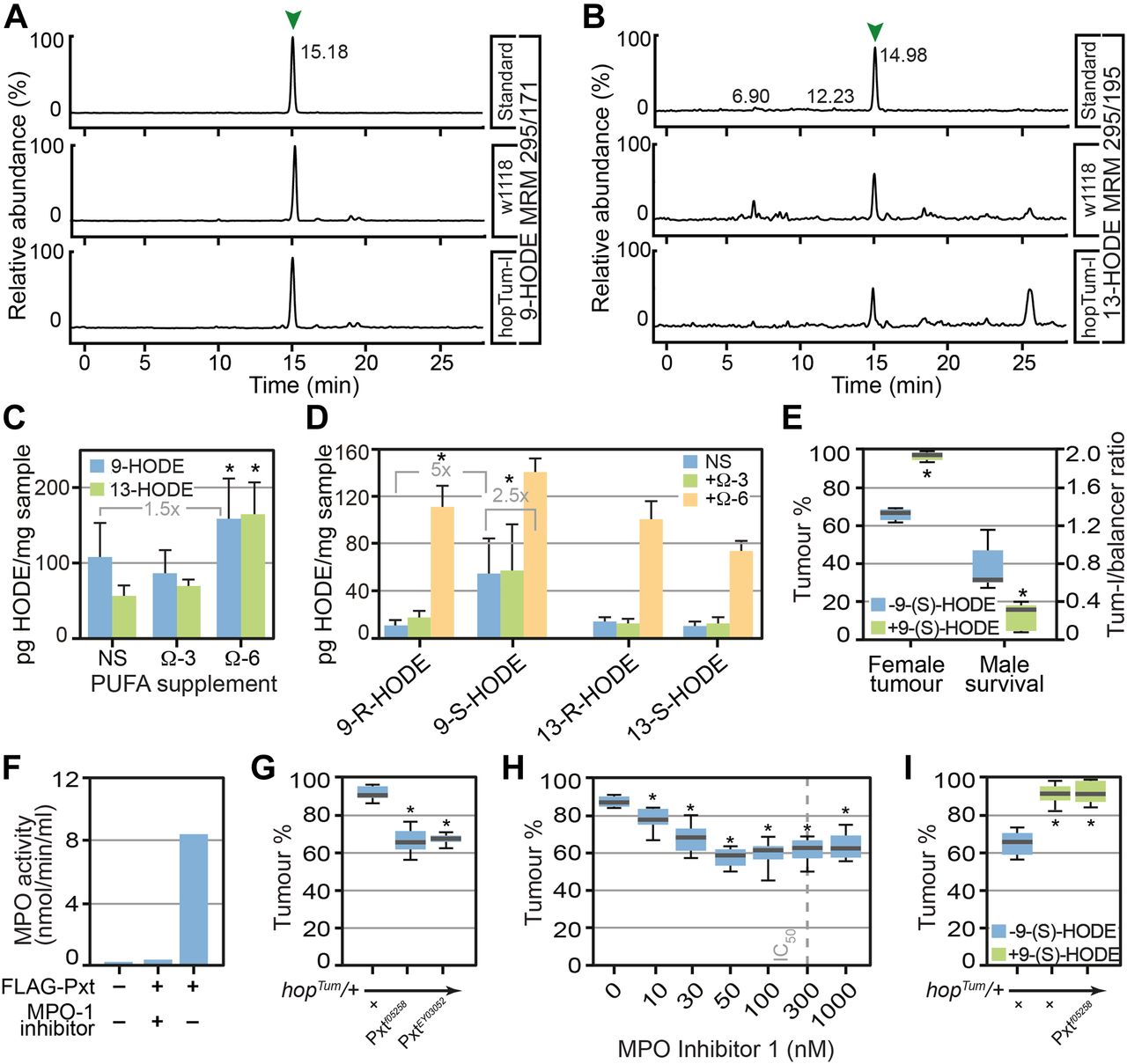
- Figure 2. Drosophila produce the lipid mediator 9-HODE which is required for inflammatory responses.
Omega-6 PUFA LA can be processed either by COX, lipoxygenase, or myeloperoxidase (MPO) enzymes to produce hydroxy-octadecadienoic acids (HODEs). (A, B) 9-HODE (MRM m/z 295>171) and (B) 13-HODE (MRM m/z 295>195) were identified in Drosophila lipid extracts. (C) The concentrations of 9-HODE and 13-HODE were unaltered after ALA (omega-3 PUFA) supplementation but increased after LA supplementation (omega-6 PUFA). Data are mean and SD of three independent determinations. (D) Chiral LC-MS/MS shows that 9-(S)-HODE was the predominant species on non-supplemented (NS) and ALA supplemented medium, whereas LA supplementation promoted the production of both 9- and 13-HODE, at (R) and (S) configuration. (E) 9-(S)-HODE (1 μM) supplementation increased melanotic tumour incidence in hopTum heterozygous females and increased lethality of hopTum heterozygous males. (F) Purified Pxt exhibited MPO activity in vitro. 3xFLAG-tagged Pxt haem peroxidase domain was over-expressed in HeLa cells, purified, and assayed for MPO activity. MPO Inhibitor-1 was used to inhibit peroxidase activity. (G) Reduction in levels of Pxt using Pxtf05258 and PxtEY03052 suppressed melanotic tumours. (H) MPO inhibitors suppressed melanotic tumours, IC50 of MPO inhibitor-1 is indicated. For (G, H), tumour incidence was assayed in heterozygous COX/MPO mutant/hopTum females raised at 27°C on medium supplemented with 0.15% LA. (I) Increased melanotic tumour incidence in hopTum heterozygous females raised on 9(S)-HODE-containing media (1 μM) was not suppressed by reduction in Pxt levels using Pxtf05258. Data information: in (C, D), three replicates were used for each assay point. * indicates values statistically significantly different from control, P-value < 0.05 determined using t test. In (E, F, G, H, I) 10 replicate crosses were used for each assay point. Box and whiskers plots were generated using R. * indicates values statistically significantly different from unsupplemented, P-value < 0.001 determined using t test. NS indicates not statistically significantly different.
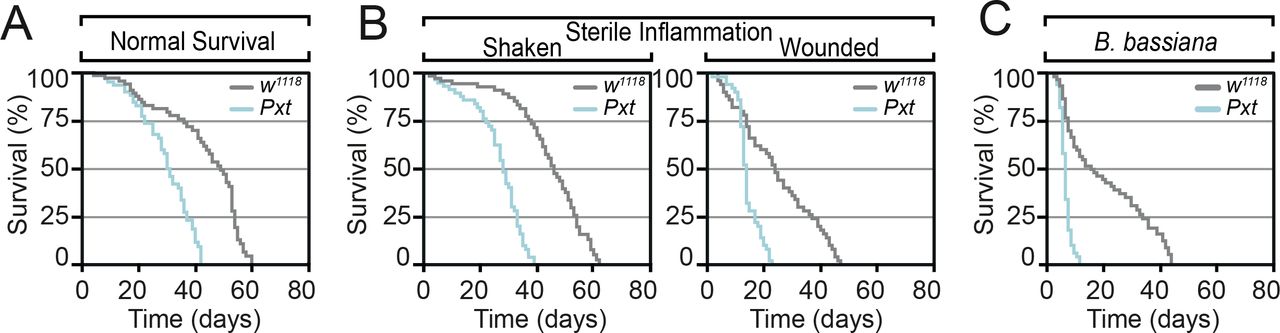
- Figure S2. Pxt is required for inflammatory responses.
(A, B, C) Pxtf05258 homozygous mutants show reduced lifespan relative to control flies (A) under control laboratory conditions, (B) after sterile wounding and (C) in response to fungal. Data information: mean survival in (A, B, C) was calculated from five cohorts of 50 flies per genotype/condition assayed every 24 h.
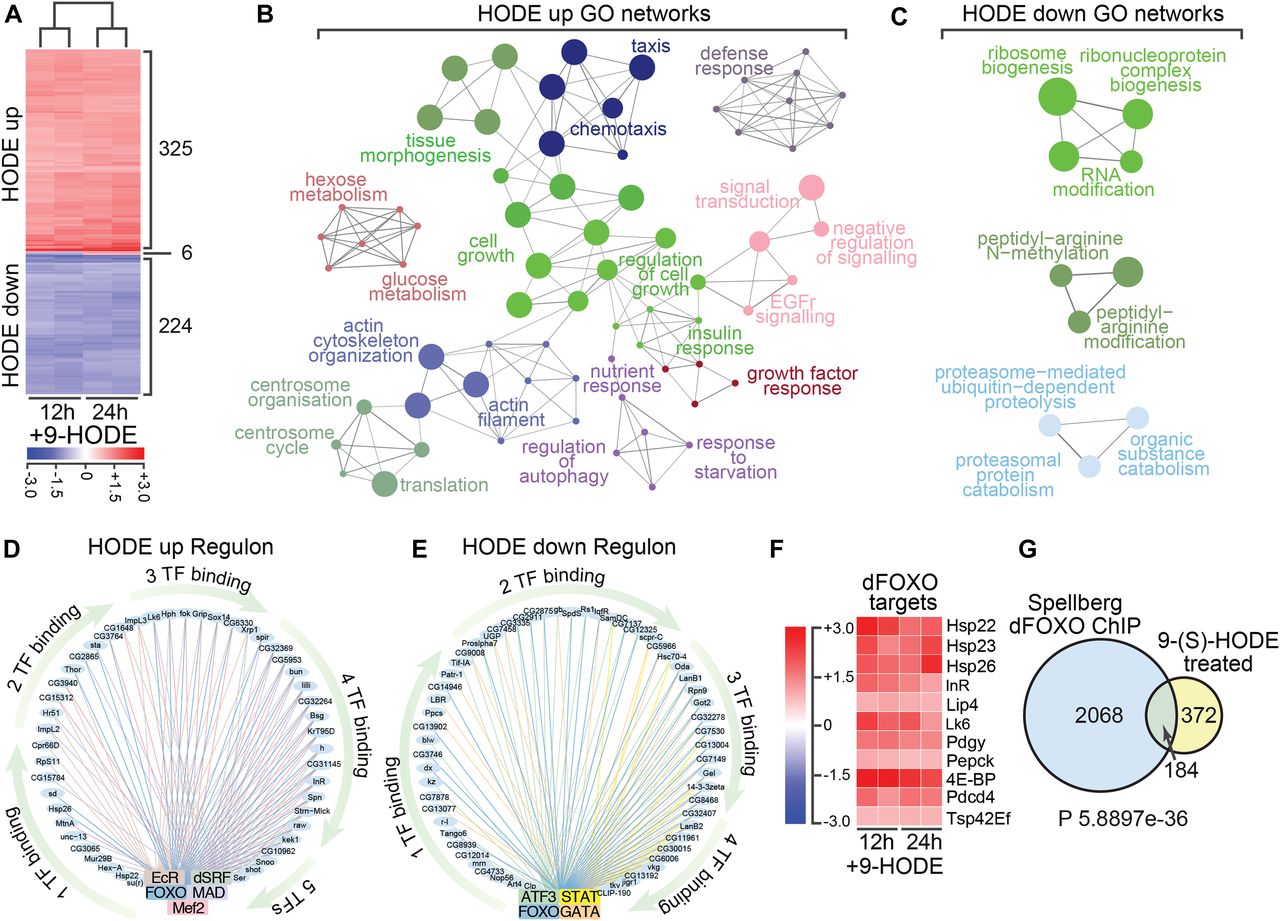
- Figure 3. Transcriptome analysis reveals 9-(S)-HODE regulates dFOXO targets.
(A) Heat map showing differentially expressed genes in S2 cells 12 and 24 h after 9-(S)-HODE treatment. Scale bar indicates fold-change relative to mock-treated control. (B) Gene ontology (GO) network analysis of the 325 genes significantly up-regulated in S2 cells 24 h after 9-(S)-HODE treatment. (C) GO network analysis of the 224 genes significantly down-regulated in S2 cells 24 h after 9-(S)-HODE treatment. (D, E) iRegulon analysis of up- and down-regulated genes defines TF regulatory circuits that transduce 9-(S)-HODE signals. Binding sites for dFOXO are a common feature of both sample sets. (F) Heat map showing fold-change in transcript levels of known dFOXO target genes after 9-(S)-HODE treatment. (G) Venn diagram showing overlap between 9-(S)-HODE transcriptional targets and genes previously defined as dFOXO targets by chromatin immunoprecipitation (ChIP) on chip analysis (Spellberg dFOXO ChIP). Hypergeometric test was performed in R to determine significance of overlap.
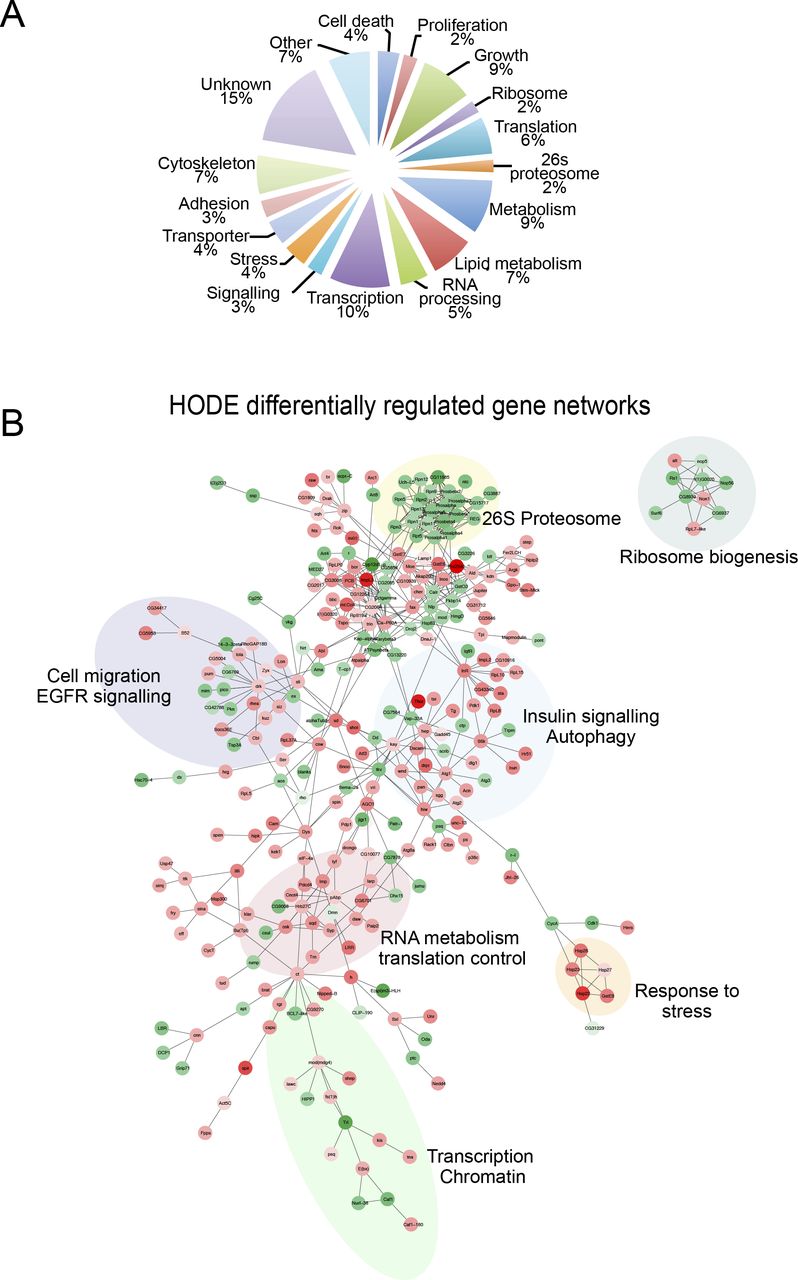
- Figure S3. 9-(S)-HODE differentially regulated networks.
(A) GO classification of 9-(S)-HODE–regulated genes. (B) Network analysis of differentially expressed genes using known Drosophila protein–protein and genetic interactions (BioGRID) or GO annotations.
- Figure 4. 9-(S)-HODE activates dFOXO.
(A) 9-(S)-HODE (10 μM) treatment inhibits Drosophila S2 cell proliferation. (B) FACS cell cycle analysis confirms cell cycle arrest. G1 and G2/M peaks seen in mock-treated S2 cells were absent in 9-(S)-HODE–treated cells replaced by a broad sub-G1 peak. (C) Western analysis of dFOXO levels in S2 cells 24 h after 9-(S)-HODE treatment. MAb E7 anti-tubulin was used as a loading control. (D) Immunofluorescence microscopy of Drosophila S2 cells shows elevated levels of nuclear dFOXO after 9-(S)-HODE treatment. (E) Quantitation of nuclear dFOXO signals in S2 cells after mock or 9-(S) HODE treatment. (F) Live imaging of 9-(S)-HODE–treated dFOXO-mCherry fat body confirming 9-(S)-HODE elevates nuclear dFOXO within 10 min of treatment. (G) Ratio of dFOXO ChIP signal at known dFOXO targets and control genomic regions in S2 cells treated with 9-(S)-HODE for 24 h relative to mock/untreated dFOXO ChIP signal. (H) Immunofluorescence microscopy of Drosophila S2 cells reveals elevated levels of nuclear dFOXO after 9-(S)-HODE treatment is resistant to insulin treatment. (I) Quantitation of nuclear dFOXO signals in mock or 9-(S) HODE treated S2 cells that are subsequently treated with insulin for 90 min before fixation. (J) 9-(S)-HODE treatment of human HeLa cells triggers nuclear entry of transiently transfected FOXO3-HA. (K) 9-(S)-HODE treatment of FOXO3-HA transfected human HeLa cells activates transcription from the FOXO3a reporter plasmid FHRE-luc. Data information: in all panels, * indicates values statistically significantly difference P-value < 0.001 determined using t test. NS, no significant difference. In (E, K), 50 determinations were used for each assay point. Box and whiskers plots were generated using R. In (G, I), three replicates were used for each assay point.
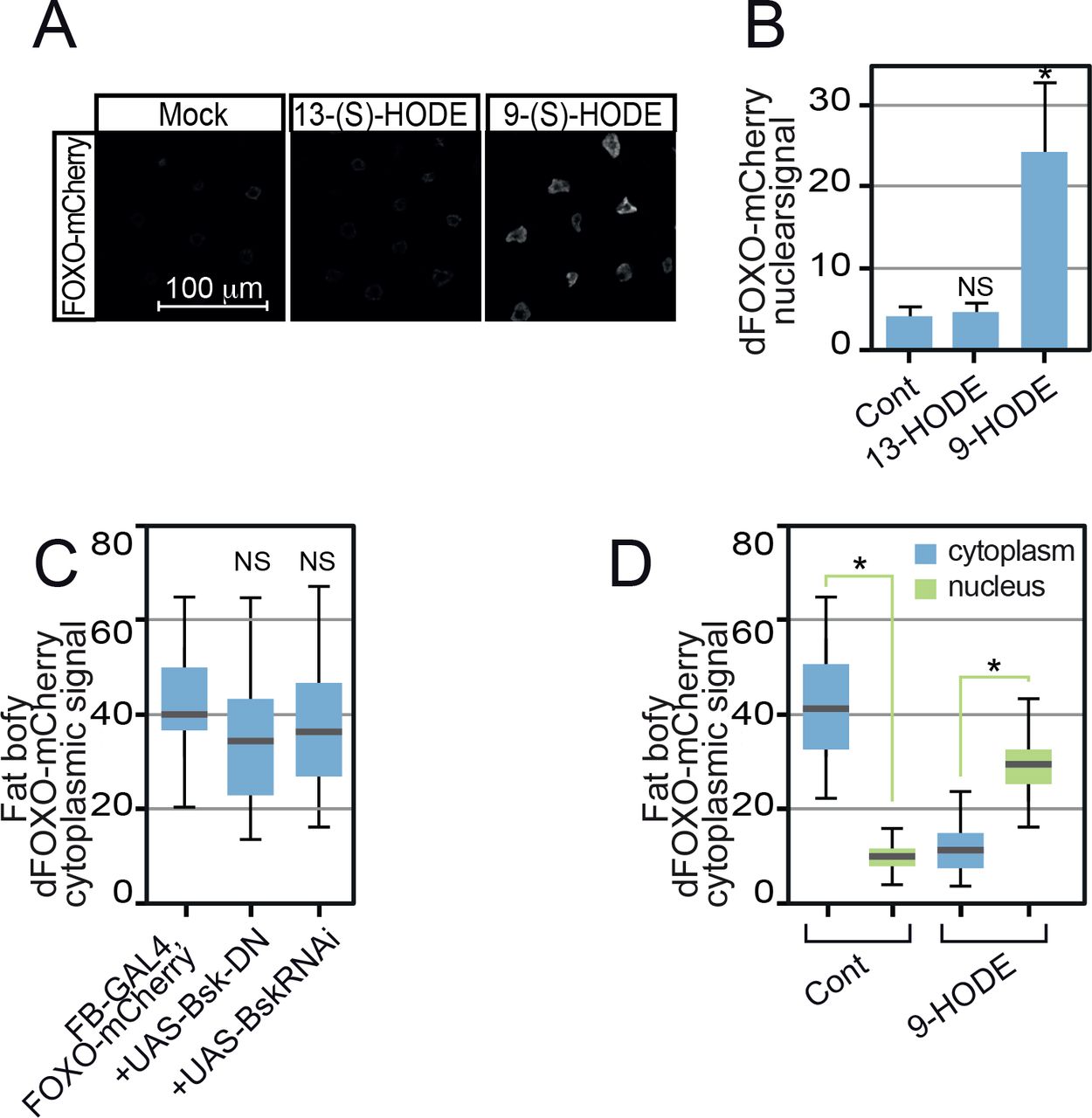
- Figure S4. 13-(S)-HODE does not activate dFOXO.
(A) Unlike 9-(S)-HODE, 13-(S)-HODE failed to induce dFOXO nuclear localisation. (B) Quantitation of dFOXO-mCherry nuclear signal. Images were collected at identical scan settings and at least 10 replicates used per data point. Data information: in (B, C, D), 50 determinations were used for each assay point. Box and whiskers plots were generated using R. * indicates values statistically significantly different from unsupplemented, P-value < 0.001 determined using t test. NS, no significant difference.
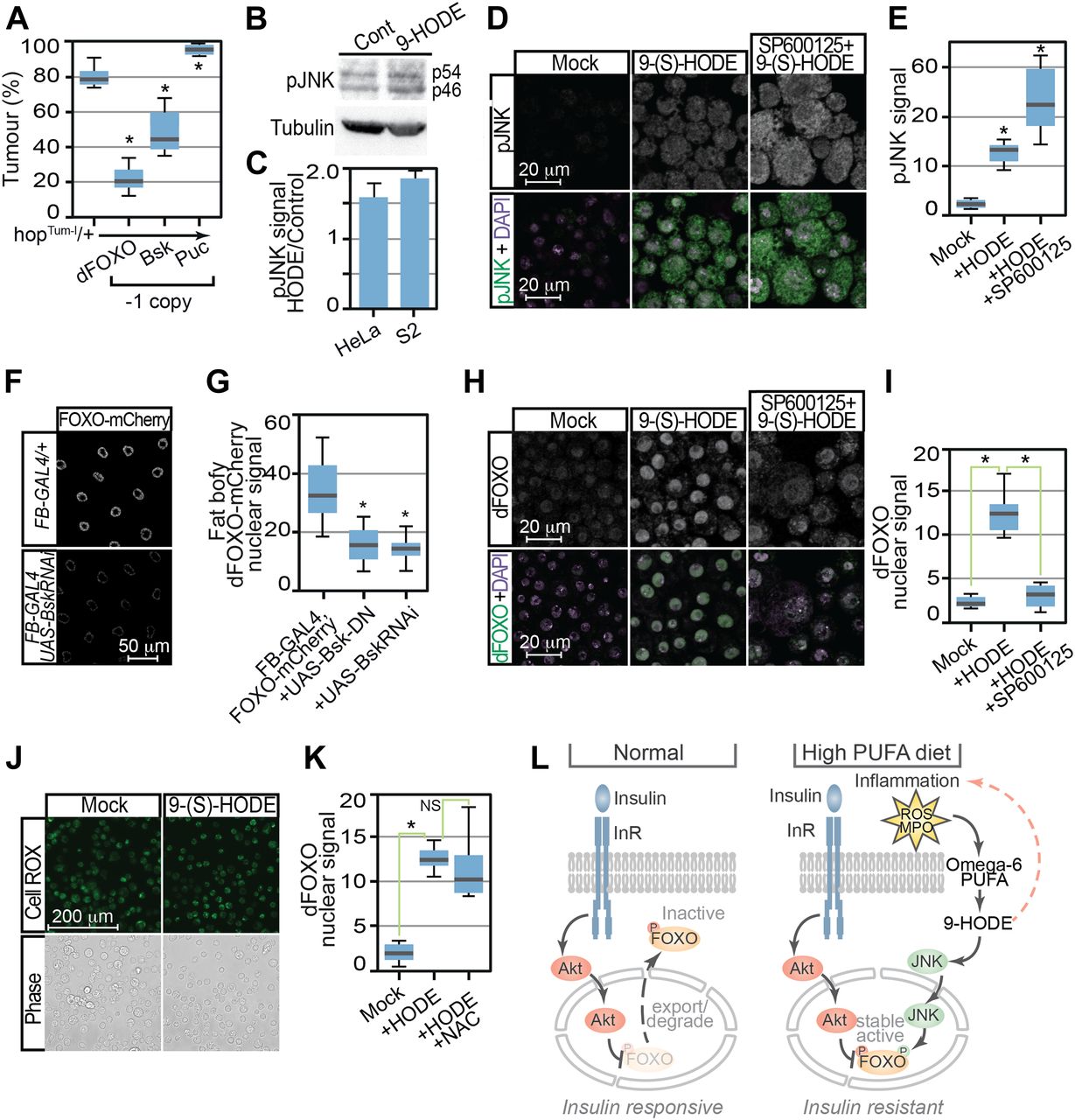
- Figure 5. Control of dFOXO by 9-HODE is mediated via the JNK pathway.
(A) Tumour incidence in heterozygous hopTum females flies was decreased by reduction in dFOXO (FoxoΔ94) or Bsk (bsk1) levels but increased by removal of Puc (pucE69). (B) JNK activation was confirmed by Western analysis of human HeLa and Drosophila S2 cells 24 h after 9-(S)-HODE treatment using anti-pJNK antibodies. MAb E7 anti-tubulin is used as a loading control. Expected pJNK (p54 and p56) species in S2 extracts are indicated. (C) Quantitation of pJNK levels in 9-(S)-HODE–treated cells relative to mock-treated control. (D) Confocal immunofluorescence microscopy of mock and 9-(S)-HODE–treated S2 cells using anti-pJNK anti-dFOXO antibodies confirms JNK activation after 9-(S)-HODE treatment. (E) Quantitation of pJNK signals in S2 cells. (F) Fat body–specific over-expression of dominant-negative Bsk variants or RNAi-mediated inhibition bsk reduces 9-(S)-HODE stimulated dFOXO-mCherry nuclear entry. (G) Quantitation of dFOXO-mCherry nuclear signal in fat body tissue. (H) Pretreatment with JNK inhibitor SP600125 prevents nuclear dFOXO localisation after 9-(S)-HODE treatment. (I) Quantitation of dFOXO nuclear signal in S2 cells after treatment with 9-(S) HODE alone or with the JNK inhibitor SP600125. (J) 9-(S) HODE treatment of S2 cells does not increase reactive oxygen species as revealed by CellROX staining. (K) Reactive oxygen species are not required for 9-(S)-HODE–triggered dFOXO nuclear localisation as simultaneous treatment with N-acetyl-L-cysteine (NAC) does not prevent nuclear entry. (L) Proposed model indicating how 9-HODE activation of FOXO could confer insulin resistance. Data information: in (A), 10 replicate crosses were used for each assay point. Box and whiskers plots were generated using R. * indicates values statistically significantly different from unsupplemented, P-value < 0.001. In (E, G, I, K), 50 determinations were used for each assay point. Box and whiskers plots were generated using R. * indicates values statistically significantly different from unsupplemented, P-value < 0.001 determined using t test.






Tables
Key Resources Table.
Reagent or resource Source Identifier Antibodies Mouse anti-B-tubulin MAb E7 Developmental Studies Hybridoma Bank RRID:AB_528499 Mouse anti-phospho-SAPK/JNK (Thr183/Tyr185) MAb G9 Cell Signalling Technology RRID:AB_2307321 Rabbit anti-dFOXO Michael Marr BRD10 Cy3-conjugated AffiniPure goat anti-rabbit antibody Jackson ImmunoResearch Cat. no. 111-167-003 FITC-conjugated AffiniPure donkey anti-mouse antibody Jackson ImmunoResearch Cat. no. 715-095-151 Anti-rabbit HRP-conjugated IgG (H+L) GE Healthcare Anti-mouse HRP-conjugated IgG (H+L) GE Healthcare Rat monoclonal anti-HA-tag [7C9] ChromoTek Deposited data Agilent microarray data This article Array express: E-MTAB-6253 Experimental models: organisms/strains Drosophila melanogaster: Schneider 2, CVCL_Z232 Invitrogen RRID: CVCL_Z232 Homo sapiens: HeLa CCL-2 American Type Culture Collection (ATCC) RRID: CVCL_0030 D. melanogaster: hopTum Bloomington Drosophila Stock Center RRID: BDSC_8492 D. melanogaster: w1118 Bloomington Drosophila Stock Center RRID: BDSC_5905 D. melanogaster: w1118; foxoΔ94/TM6B, Tb1 Bloomington Drosophila Stock Center RRID: BDSC_42220 D. melanogaster: w1118 P{w+mC=UAS-bsk.DN}2 Bloomington Drosophila Stock Center RRID: BDSC_6409 D. melanogaster: bsk1 cn1 bw1 sp1/CyO, Bloomington Drosophila Stock Center RRID: BDSC_3088 D. melanogaster: w*; cno3 P{ry+t7.2=A92}pucE69/TM6B, P{w+mC=iab-2(1.7)lacZ}6B, Tb1 Bloomington Drosophila Stock Center RRID: DGGR_109029 D. melanogaster: w1118; P{GD10555}v3413 UAS-bskRNAi Bloomington Drosophila Stock Center RRID:FlyBase_FBst0460476 D. melanogaster: w1118; P{GAL4}fat (FB-GAL4) Gronke et al (2003) RRID:FlyBase_FBti0013267 D. melanogaster: w1118; P{GD10555}v3413 UAS-bskRNAi Bloomington Drosophila Stock Center RRID:FlyBase_FBst0460476 D. melanogaster: y1 w67c23; P{EPgy2}PxtEY03052 Bloomington Drosophila Stock Center RRID:BDSC_15620 D. melanogaster: PBac{WH}Pxtf05258 The Exelixis Collection at the Harvard Medical School RRID: FBti0067872 D. melanogaster: dfoxo-v3-mCherry Kakanj et al (2016) N/A Beauveria bassiana CABI (T. Petch, 1947) IMI: 12942 Oligonucleotides foxo-ChIP-control forward primer (5′-AGCCCGCGAAGATACAAGAG-3′) This article N/A foxo-ChIP-control reverse primer (5′-AGCCACAAACAGCGACAGAA-3′) This article N/A pdcd4-ChIP forward primer (5′-CCGTTGGGAGTCTCTCTCTC-3′) This article N/A pdcd4-ChIP reverse primer (5′-GCTCTCCCCACCTTCTCACC-3′) This article N/A hsp26-ChIP forward primer (5′-GTGCGCCTGTATGAGTGAGA-3′) This article N/A hsp26-ChIP reverse primer (5′-GTGGGAGATTGCTGGCGTTA-3′) This article N/A 4EBP-ChIP forward primer (5′-CAAGAACCAGCCGGTTTGTC-3′) This article N/A 4EBP-ChIP reverse primer (5′-GCTCTCTTCTCGCTCTTTCG-3′) This article N/A Pxt-RT-PCR forward primer (5′-GCAGCTCCTCGATGTGATTGAAAC-3′) This article N/A Pxt-RT-PCR reverse primer (5′-CTAGAGTGCGAGCGAGAGGTAAGA-3′) This article N/A RpL32-RT-PCR forward primer (5′-ATCCGCCACCAGTCGCATCGATATGCTAAG-3′) This article N/A RpL32-RT-PCR reverse primer (5′-TCTTGAGAACGCAGGCGACCGTTGGGGTTG-3′) This article N/A DptA-RT-PCR forward primer (5′-GCAGTTCACCATTGCCGTCGCCTTACTTTG-3′) This article N/A DptA-RT-PCR reverse primer (5′-TGAAGATTGAGTGGGTACTGCGGTGGTGGA-3′) This article N/A Drs-RT-PCR forward primer (5′-ATCAAGTACTTGTTCGCCCTCTTCGCTGTC-3′) This article N/A Drs-RT-PCR reverse primer (5′-CTCGTTGTCCCAGACGGCACAGGGACCCTT-3′) This article N/A Recombinant DNA Plasmid: pTRE3G Clontech 631173 pTRE3G-3XFLAG This article N/A pTRE3G-3XFLAG-Pxn This article N/A Software and algorithms FIJI Schindelin et al (2012) https://imagej.net/Fiji Marray Paquet (2009) https://bioconductor.org/packages/release/bioc/html/marray.html Limma Ritchie et al (2015) http://bioconductor.org/packages/release/bioc/html/limma.html Bioconductor version 3.4 http://bioconductor.org https://bioconductor.org/news/bioc_3_4_release/ R version 3.1.2 https://www.R-project.org https://cran.r-project.org/bin/windows/base/old/3.1.2/ ClueGo (Bindea et al, 2009) https://apps.cytoscape.org/apps/cluego esyN (Bean et al, 2014) http://www.esyn.org Cytoscape (Shannon et al, 2003) https://cytoscape.org iRegulon (Janky et al, 2014) http://iregulon.aertslab.org
Supplemental Data 1.
Gene with most significant changes in expression following 12 h 9-(S)-HODE treatment.[LSA-2019-00356_Supplemental_Data_1.xlsx]
Supplemental Data 2.
Gene with most significant changes in expression following 24 h 9-(S)-HODE treatment.[LSA-2019-00356_Supplemental_Data_2.xlsx]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Related Articles
Cited By...
- No citing articles found.