




Article Figures & Data
Figures
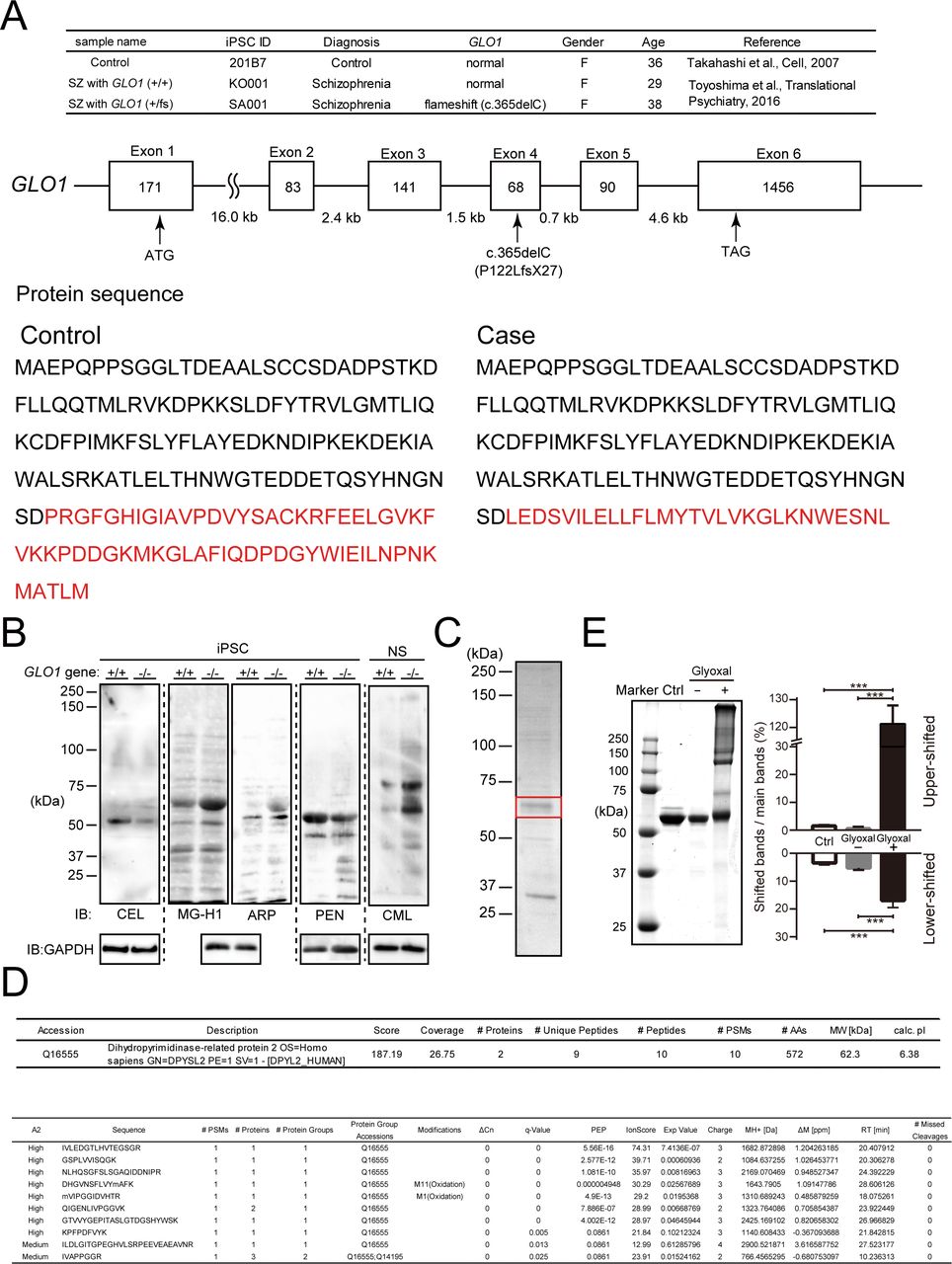
- Figure S1. Genetic background of human iPS cells and identification of AGE-modified proteins in GLO1 ( − / − ) iPS cells.
(A) Genetic background of human iPS cells. iPS cells were derived from a schizophrenia (SZ) patient with the GLO1 frameshift mutation (SZ+GLO1 (fs)), a case–control schizophrenia patient with copy number variation (CNV) of the 22q11.2 deletion without a frameshift mutation (SZ) and a healthy control (control). The frameshift mutation (c.365del, P122LfsX27) is located within the fourth exon of the GLO1 gene, and the divergence of the GLO1 protein sequence resulting from the frameshift mutation is shown at the bottom panel. (B) Immunoblotting of AGE-modified proteins in GLO1 (−/−) iPS cells by anti-AGE antibodies that detect N-ε-(carboxymethyl) lysine (CML), N-ε-(carboxyethyl) lysine (CEL), methylglyoxal-derived hydroimidazolone (MG-H1), argpyrimidine (ARP), and PEN. (C) CBB-stained SDS–PAGE gels of AGE-proteins containing the 60-kD CML protein. The 60-kD band (boxed in red) was cut off for protein identification by MS/MS analysis. (D) Identification of the major CML-modified protein by LC-MS/MS as CRMP2 (DPYSL2). (E) In vitro glyoxal-induced AGE-modified recombinant CRMP2 proteins. The samples of nonincubated CRMP2 protein and CRMP2 protein incubated with or without glyoxal were assessed by SDS–PAGE shown in the left panel, and the quantification analysis is shown in the right panel (statistics: the bar graph represents the means ± SDs; ***P < 0.001, as determined by one-way ANOVA followed by Tukey’s multiple-comparisons test; n = 3).
Source data are available for this figure.
Source Data for Figure S1[LSA-2019-00478_SdataFS1.pdf]
- Figure 1. iPS cells from a schizophrenia patient with a GLO1 mutation exhibited impaired neurosphere growth under enhanced carbonyl stress, and CRMP2 is a major target of AGE modification under enhanced carbonyl stress in GLO1 (−/−) and GLO1 (fs) iPS cells.
(A) Bright-field images of neurospheres generated from iPS cells derived from a control individual, a schizophrenia (SZ) patient with wild type GLO1 gene (SZ with GLO1 (+/+)), and an SZ patient with the GLO1 frameshift mutation (SZ with GLO1 (fs)) (scale bar: 200 μm). (B) Quantification of the number of neurospheres. (C) Immunoblotting of GLO1 protein in GLO1 (+/+) and GLO1 (−/−) iPS cells. (D) Bright-field images of neurospheres that formed from GLO1 (−/−) iPS cells in the presence or absence of pyridoxamine (scale bar: 200 μm). (E, F) Quantification of the size (E) and number (F) of neurospheres from GLO1 (+/+) and GLO1 (−/−) iPS cells. (G) Phase-contrast images of neurons derived from GLO1 (+/+) and GLO1 (−/−) iPS cells in the presence or absence of pyridoxamine (scale bar: 25 μm). (H) Quantification of the neurite length from images shown in (G) (statistics: data are presented as the means ± SDs; *P < 0.05, **P < 0.01, and ***P < 0.001, as determined by two-way ANOVA followed by Tukey’s multiple-comparisons test; (B), (E), and (F): n = 60; (H): n = 30). (I) Immunoblotting of AGE-modified proteins in GLO1 (−/−) iPS cells by anti-AGE antibodies that detect N-ε-(carboxymethyl) lysine (CML). (J) Immunoblotting of the CML modification at ∼60 kD was gradually decreased by pyridoxamine treatment in a dose-dependent manner. (K) Quantification of CML modification at ∼60 kD after pyridoxamine treatment (statistics: data are presented as the means ± SDs; **P < 0.01 and ***P < 0.001, as determined by one-way ANOVA followed Tukey’s multiple comparison test; n = 9). (L) CML-modification of CRMP2 immunoprecipitated by anti-CRMP2 antibody in GLO1 (−/−) and GLO1 (fs) iPS cells. IB, immunoblot; IP, immunoprecipitate.
Source data are available for this figure.
Source Data for Figure 1[LSA-2019-00478_SdataF1.pdf]
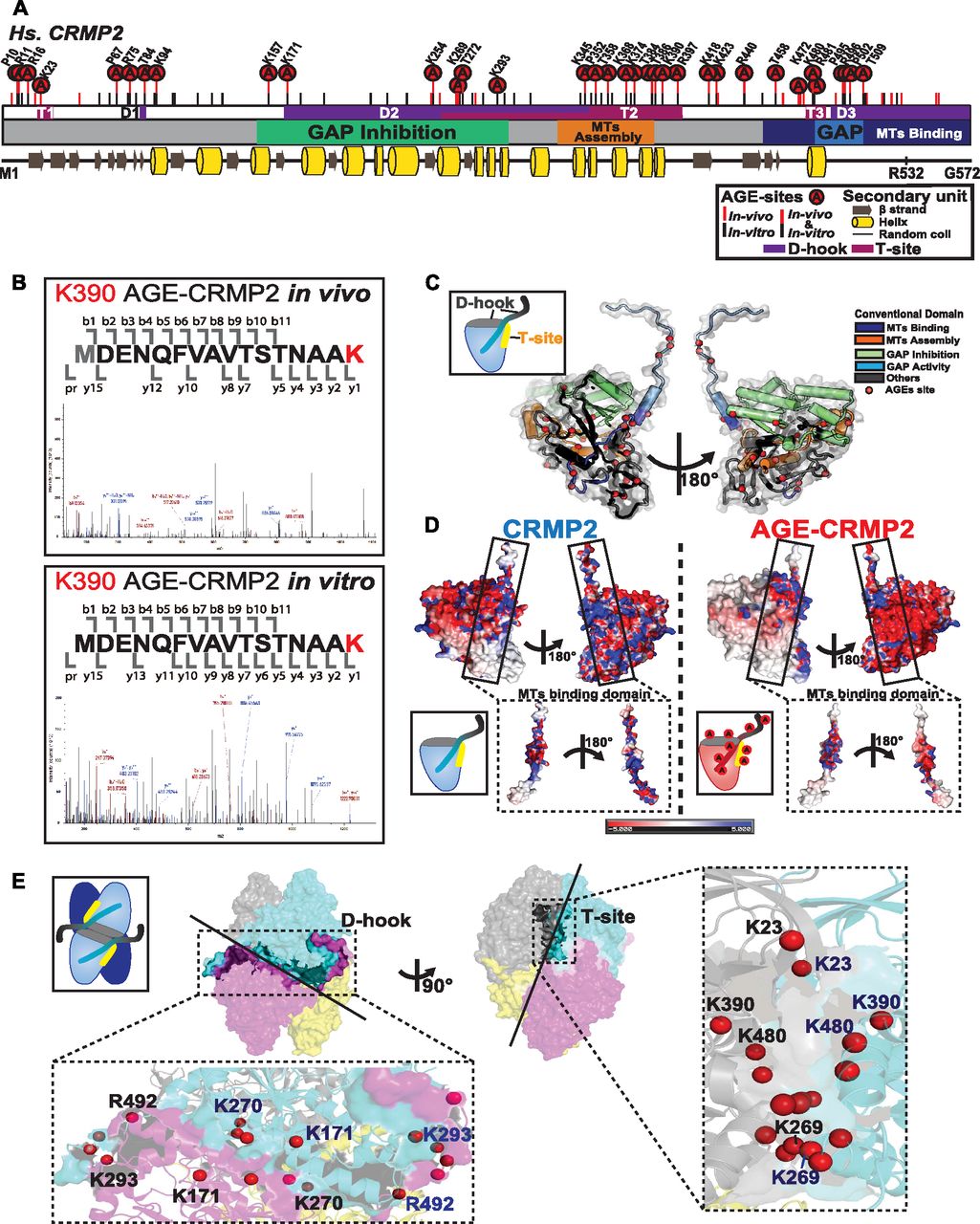
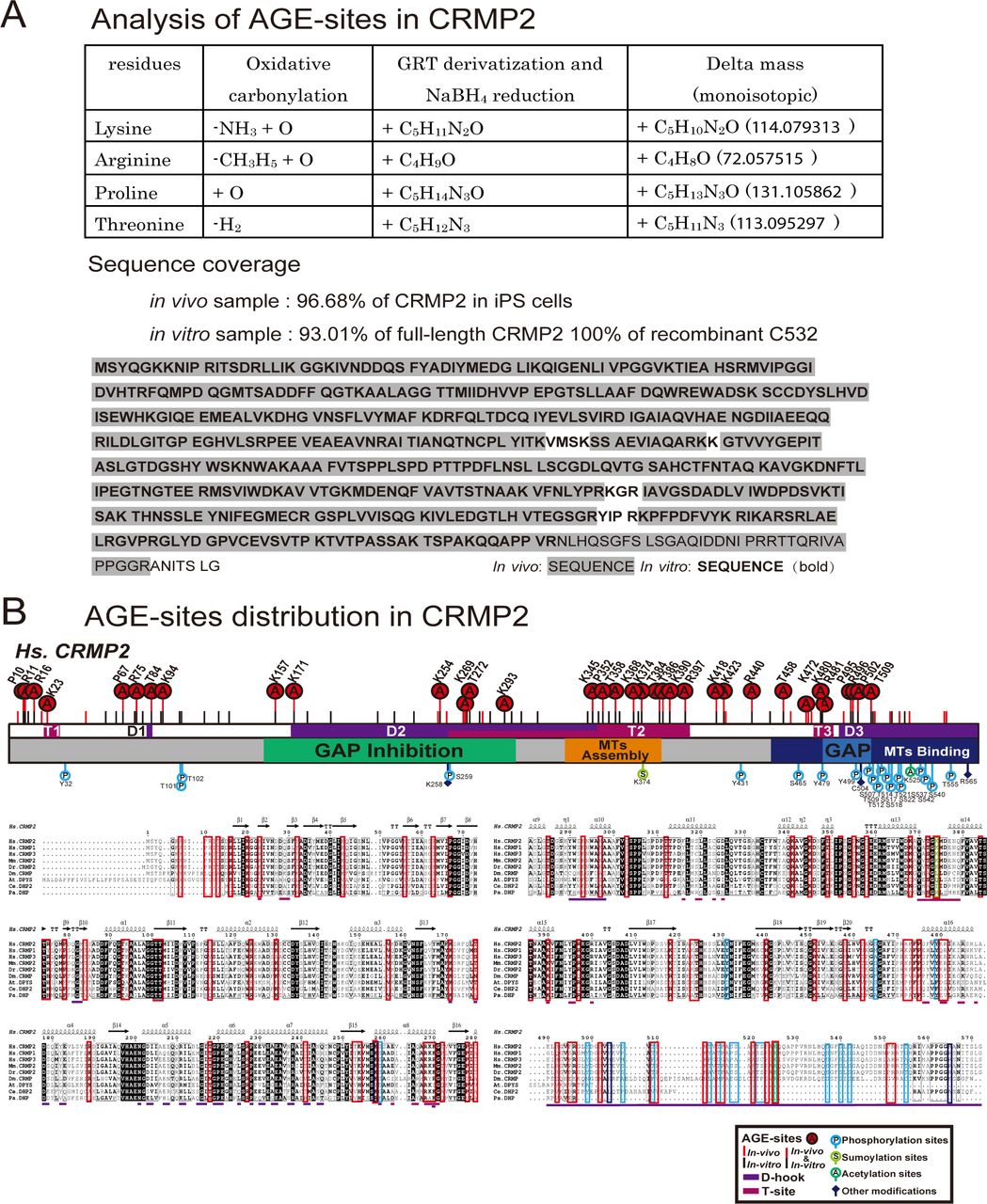
- Figure 2. AGE sites are widely distributed around conventional functional domains and located in two structural interfaces of the functional tetramer complex, D-hook and T-site.
(A) Distribution of AGE sites in CRMP2 in vivo and in vitro. 29 (7 Arg, 11 Lys, 5 Pro, and 6 Thr) overlapped (indicated by “A” in a red circle) among the 50 sites in in vivo CRMP2 C532 (showed with a red line) and 62 sites in in vitro CRMP2 C532 (shown with a black line). The conventional motifs identified in previous studies related to MT assembly are shown at the top, and the 2D structural units are shown at the bottom. (B) An AGE site on K390 was determined with high accuracy based on the MS/MS spectra obtained both in vivo and in vitro. (C) AGE sites are distributed on the outer surface around the conventional functional domains in the CRMP2 structure: GAP activity (6 sites), inhibition of GAP activity (19 sites), promotion of MT assembly (10 sites), and interaction with MTs (21 sites). The structure model (PDB code: 5MKV) was used for representation here. (D) Simulation of surface potential changes of CRMP2, which showed that AGE modification induced a great loss of positive charges (in blue) and an apparent increase in negative charges (in red). The MT-binding domain is highlighted. The structure models (PDB codes: 6JV9 and 6JVB) were used for simulation analysis. (E) AGE sites were localized in two structural interfaces associated with the self-assembly of CRMP2: D-hook and T-site. D-hook comprises 10 and 8 AGE sites for molecule A (in magenta) and the adjacent molecule B (in light blue). In total, 18 AGE sites of D-hook are shown in the bottom-left panel. T-site contains 9 AGE sites for each molecule. In total, 18 AGE sites of T-site are shown in the right panel. Representative AGE-modified residues are labelled.
- Figure S2. Determination of AGE sites in both the in vivo CRMP2 protein purified from GLO1-KO iPS cells and recombinant AGE-CRMP2 protein in vitro by LC-MS/MS analysis.
(A) Advanced proteomic strategy for detection of the AGE modification. The derivatization of AGE sites by GRT labelling produces the additional mass on the AGE-modified peptides, which is detected in the m/z spectrum (upper panel). The sequence coverage of the LC-MS/MS analyses is also shown (bottom panel). The detected peptides of in vivo and in vitro CRMP2 are indicated with grey shade and bold font, respectively. (B) Distribution of AGE sites and other reported PTMs on functional domains and structural interfaces (D-hook and T-site) of CRMP2.
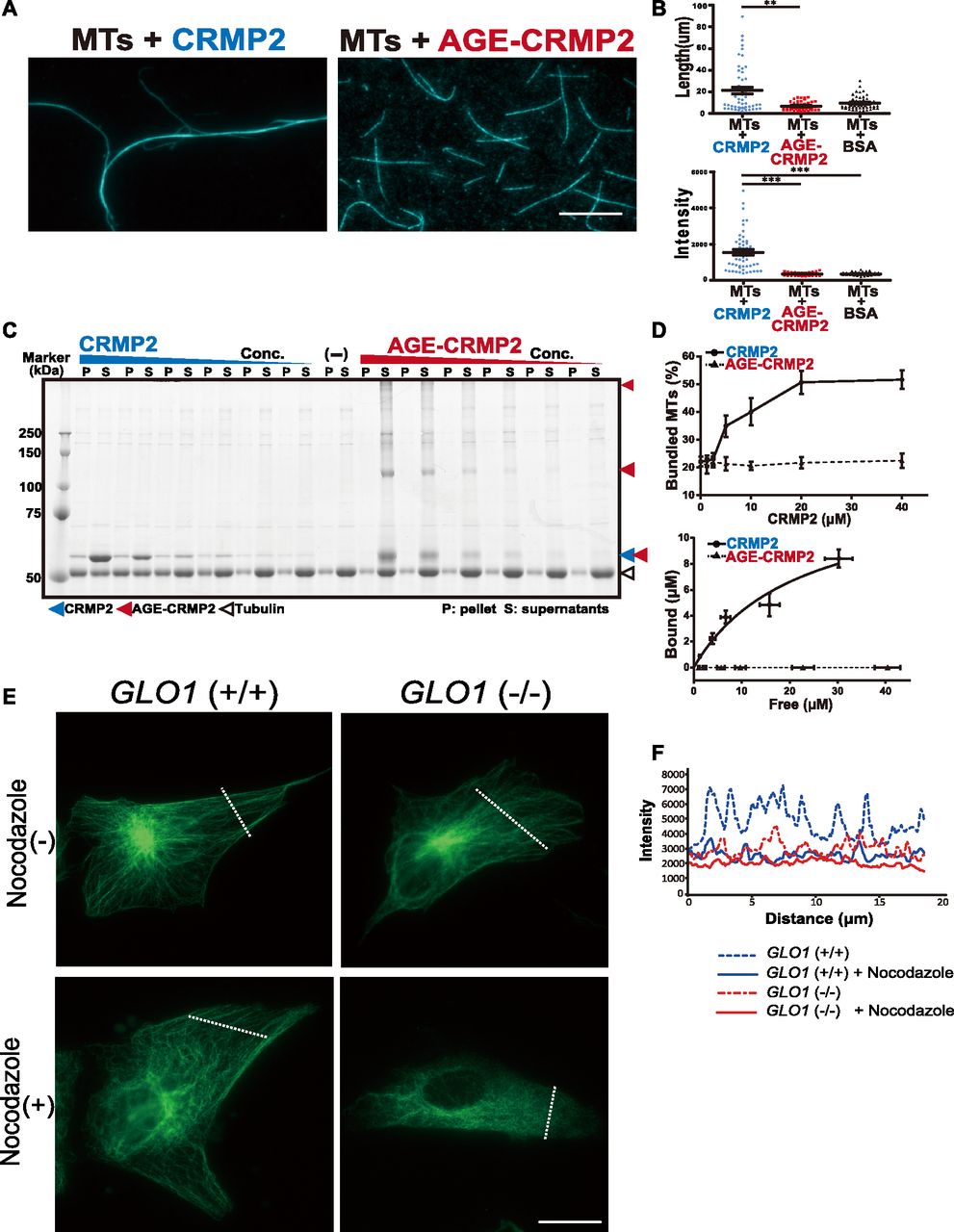
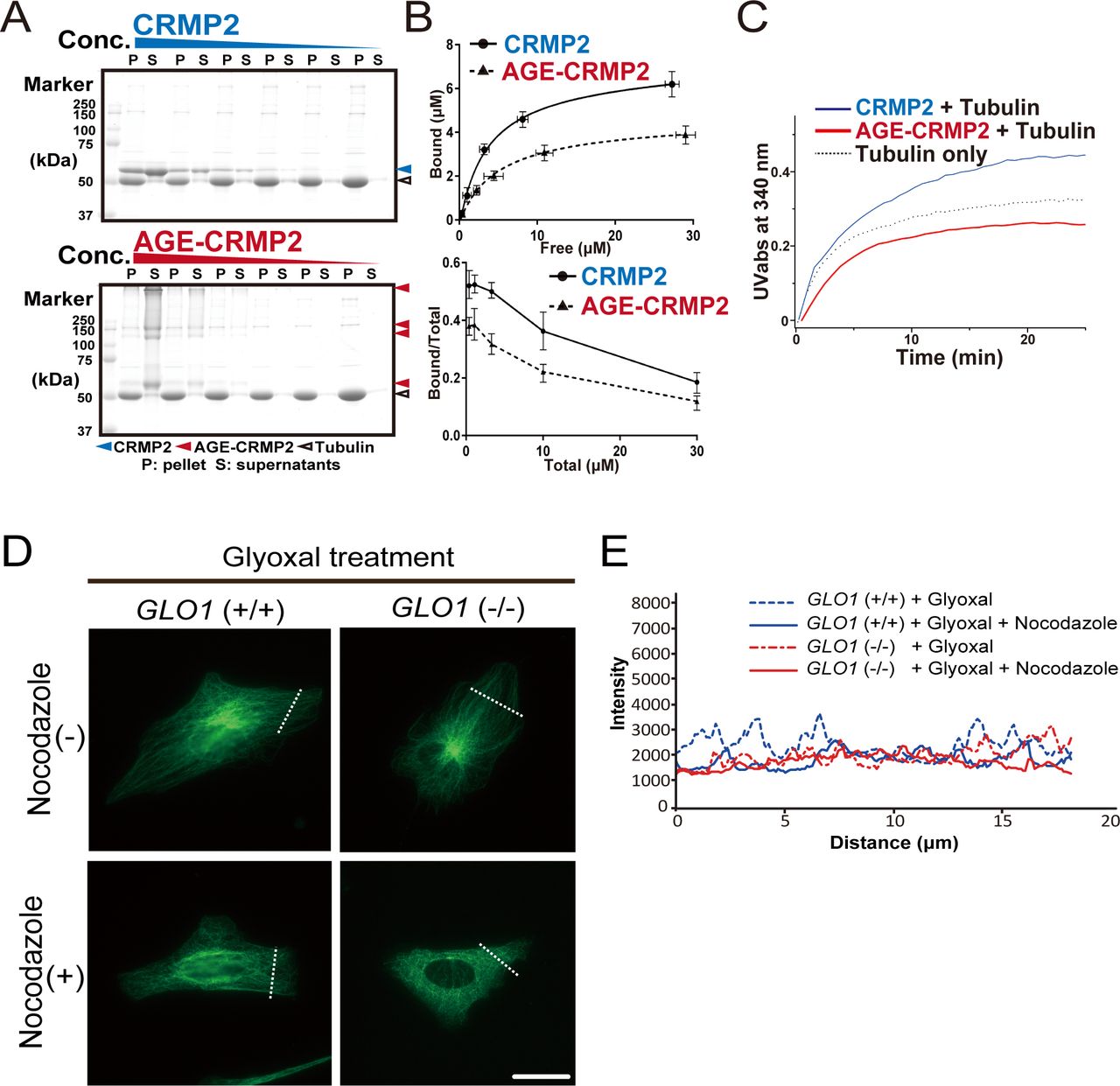
- Figure 3. AGE modification diminishes the MT-bundling activity of CRMP2.
(A) CRMP2 C532 protein significantly bundled polymerized MTs (protein ratio = 1 μM tubulin dimer: 4 μM CRMP2, left panel). In contrast, AGE-CRMP2 did not display MT-bundling activity (protein ratio = 1 μM tubulin dimer: 4 μM AGE-CRMP2, right panel) (scale bar: 10 μm). (B) Quantification of the length and intensity of MTs (statistics: data are presented as the means ± SEMs; **P < 0.01 and ***P < 0.001, as determined by the Kruskal–Wallis with Dunn’s multiple comparison test; n = 50). (C) Low-speed MT co-sedimentation assay for assessing the bundling activity of CRMP2 (or AGE-CRMP2) with polymerized MTs. Various concentrations of proteins were mixed with Taxol-stabilized MTs and then subjected to co-sedimentation (6,000g, 10 min) assessment, and the bundled MTs were then sedimented to obtain a pellet (P). The P and supernatant (S) fractions were used to assay the MT-bundling and MT-binding activities of unmodified and AGE-modified CRMP2 to bundled MTs. The irreversible polymers of AGE-CRMP2 were also observed as the upper bands on an SDS–PAGE gel. (D) Quantitative analysis of the low-speed co-sedimentation assay results. Three replicates of the co-sedimentation data were used for the quantification. The MTs in the P and S fractions were quantified to analyze the proportion of bundled MTs (upper panel), and the amounts of bound proteins were plotted as functions of the amounts of unbound proteins (lower panel; the bar graph represents the means ± SDs). (E) Assay of the stability of MTs in GLO1+/+ and GLO1−/− iPS cells in the presence of nocodazole, an MT-destabilizing drug (scale bar: 25 μm). (F) Comparison of the fluorescence intensity of stabilized MTs. The fluorescence intensity of MTs was measured at a distance equal to one-third of the distance from the leading edge of the cell to the edge of the nucleus.
Source data are available for this figure.
Source Data for Figure 3[LSA-2019-00478_SdataF3.pdf]
- Figure S3. Effect of glyoxal on CRMP2 activity both in vitro and in vivo.
(A) High-speed MT co-sedimentation assay to assess the total binding activity of CRMP2 (or AGE-CRMP2) with polymerized MTs. Various concentrations of CRMP2 proteins were mixed with Taxol-stabilized MTs and then subjected to co-sedimentation (76,000g, 30 min) assessment. The pellet (P) and supernatant (S) fractions were analyzed to assay the binding of unmodified and AGE-CRMP2 to MTs. (B) Quantification of high-speed co-sedimentation assay results. Three replicates of the co-sedimentation data were used for the quantification. The MT-bound proteins in the P and the unbound proteins in the S were quantified to plot both the binding affinity (upper panel) and the bound ratio (lower panel) of unmodified and AGE-modified CRMP2 (the bar graph represents the means ± SDs). (C) High-speed MT turbidity assays to compare the total activities of unmodified and AGE-modified CRMP2 to promote MT polymerization and bundling. (D) Assay of MT stability in GLO1 (+/+) and GLO1 (−/−) iPS cells under glyoxal treatment in the presence of nocodazole, an MT-destabilizing drug. (scale bar: 50 μm). (E) Comparison of the fluorescence intensity of stabilized MTs.
Source data are available for this figure.
Source Data for Figure S3[LSA-2019-00478_SdataFS3.pdf]
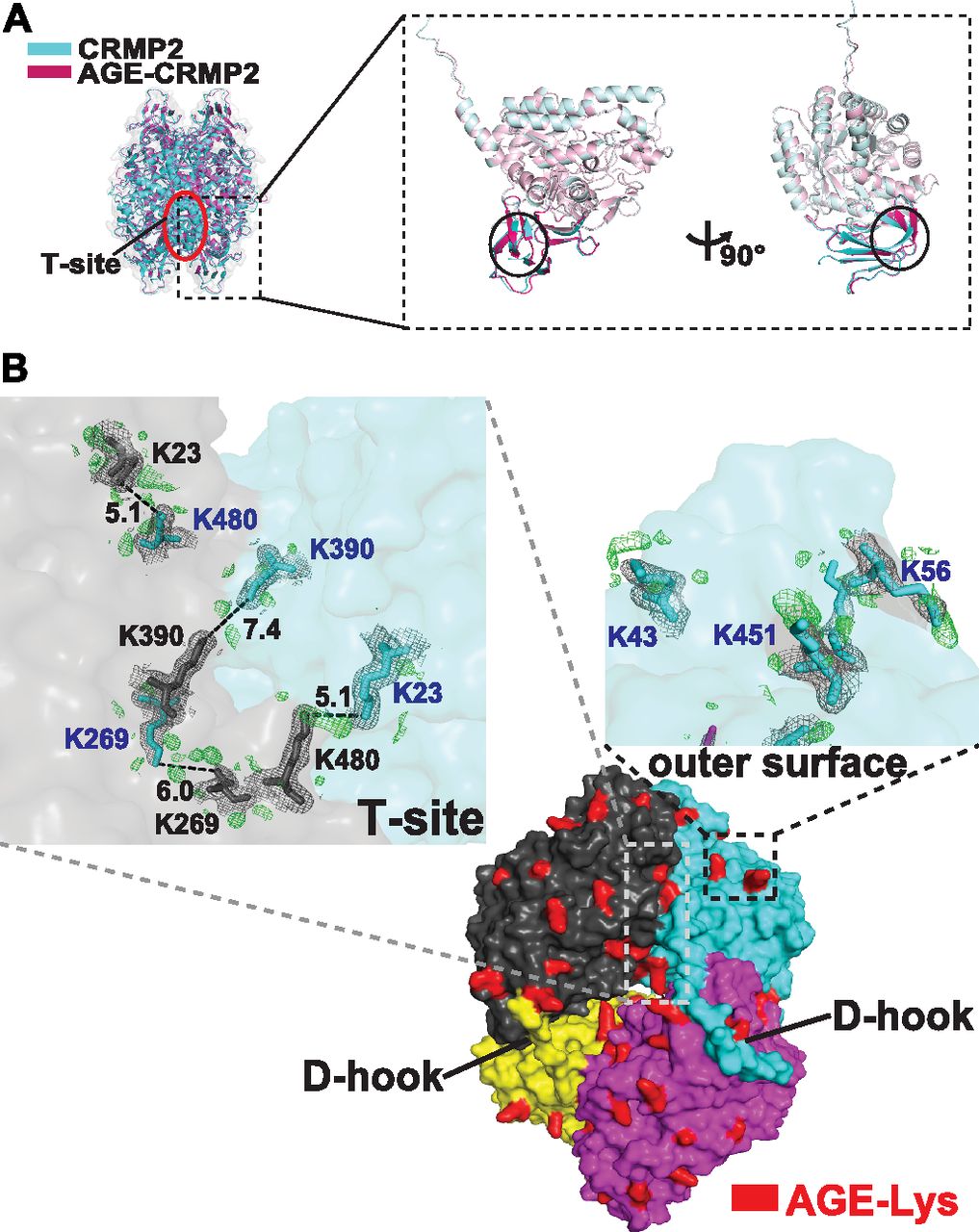
- Figure 4. AGE modification induces the formation of irreversible di-Lys bonds on T-site and the outer surface of the tetramer complex.
(A) Superposition of the tetrameric complexed structures of CRMP2 (C532, in blue) and AGE-CRMP2 (in red) solved by X-ray crystallography at 2.26- and 2.00-Å resolution, respectively. In the AGE-modified structure, the β2 sheet underwent a sharp inclination, and the β3 sheet broke into two separated β strands with a linking loop. (B) Comparison of the electron density maps of unmodified and AGE-modified CRMP2 structures. The additional electron density in the Fo-Fc map (AGE-Lys, in red-coloured density) was observed within the T-site and on the outer surface of the tetrameric complex. In the AGE–CRMP2 complex structure, three Lys pairs (K23–K480, K269–K269, and K390–K390, in the upper left panel) within the T-site and in close proximity to the ε-ammonium group were modified by AGE to likely form di-Lys bonds. Three Lys residues (K43, K56, and K451, in the upper right panel) were modified by AGE, and their modified side chains were located in outward protrusions. The 2Fo-Fc (in grey) and Fo-Fc (in green) density maps were contoured at 1σ and 3σ, respectively.
- Figure 5. AGE modification induces CRMP2 dysfunction through formation of an irreversible inactive conformation.
(A) DSC analysis of unmodified and AGE-modified CRMP2 proteins to evaluate the flexibility and dynamic size of the complexes in solution. The thermal transition midpoint (Tm) of AGE-CRMP2 (in red) increased by 1.8°C–64.4°C from that of CRMP2 (62.6°C). (B) High-resolution size exclusion chromatography (HiRes-SEC) analysis of unmodified CRMP2 (in blue) and AGE-CRMP2 (in red) proteins to evaluate the dynamic size of the complexes in solution. The chromatogram of a standard mixture of molecular weight marker proteins is displayed with a black dotted line (ferritin, 440 kD; aldolase, 158 kD; ovalbumin, 44 kD; and ribonuclease A, 13.7 kD). The irreversibility of the complex in each fraction was monitored on the SDS–PAGE with a strong reducing environment (CRMP2 in a blue box, AGE-CRMP2 in a red box). The irreversible polymers of AGE-CRMP2 were observed as the upper bands on the SDS–PAGE gel (shown in a red box). The supernatants (S, samples for HiRes-SEC) and pellets (P) of the total protein samples were also analyzed by SDS–PAGE (shown at the far left of the SDS–PAGE images) to assess the AGE-induced aggregation. (C) Immunoblotting of transfected Myc-CRMP2 in GLO1 (+/+) and GLO1 (−/−) iPS cells with or without pyridoxamine treatment. The normal bands at 60 kD (black arrow), upper shifted bands (red arrow heads), and lower shifted bands (white arrow heads) of CRMP2 are indicated. (D) Comparison with upper or lower shifted bands to normal CRMP2 band. Ratio of Upper shifted bands were significantly increased in GLO1 (−/−) iPS cells (statistics: data are presented as the means ± SDs; *P < 0.05, and ***P < 0.001, as determined by one-way ANOVA followed by Tukey’s multiple-comparisons test; n = 3).
Source data are available for this figure.
Source Data for Figure 5[LSA-2019-00478_SdataF5.pdf]
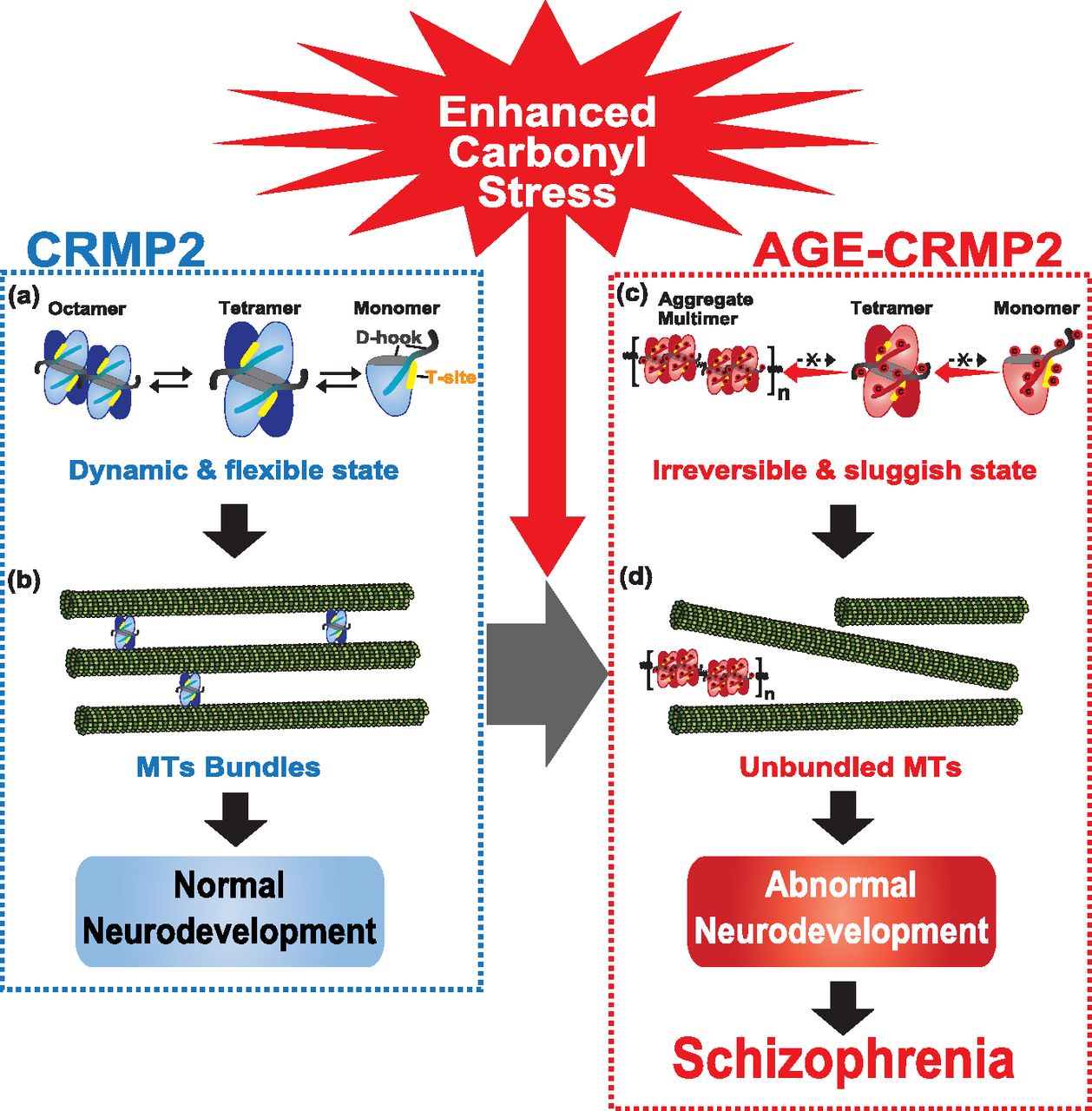
- Figure 6. Model of the molecular pathogenesis of a subset of schizophrenia under enhanced carbonyl stress.
(A) Under healthy conditions, CRMP2 proteins exist as a tetrameric complex in dynamic equilibrium between its transformative units, such as its octamer, tetramer, and monomer forms. (B) In this state, CRMP2 showed high MT-bundling activity. (C) Under enhanced carbonyl stress, CRMP2 proteins are hyper-carbonylated as the major target of AGE modification, and this effect induces the irreversible polymerization and aggregation of CRMP2 through the formation of di-Lys bonds within and between the tetrameric complexes. (D) Loss of the transformative dynamics of CRMP2 diminishes the MT-bundling activity of CRMP2, which results in abnormal cellular and developmental deficits, and this effect constitutes the pathogenic basis of a schizophrenia subset under enhanced carbonyl stress.






Tables
Unmodified C532 AGE-C532 Data collection statistics X-ray source PF-BL17A PF-BL5A Space group P21 P21 Unit cell parameters a, b, c (Å) 80.09, 158.43, 88.00 95.21, 147.33, 95.23 α, β, γ (°) 90.00, 94.19, 90.00 90.00, 103.91, 90.00 Wavelength (Å) 1.0 1.0 Resolution limits (Å)a 50.00–2.26 (2.30–2.26) 50.00–2.00 (2.06–2.00) No. of unique reflections 102,278 171,234 Completeness (%) 99.9 (99.9) 100 (100) Redundancy 3.3 (3.3) 3.8 (3.9) Rmerge (%)b 16.3 (57.9) 10.4 (62.2) Rp.i.m (%) 10.6 (37.5) 6.2 (36.3) Mean I/σ (I) 6.5 (2.5) 9.1 (1.5) CC1/2 0.96 (0.64) 0.98 (0.71) Refinement statistics Resolution limits (Å) 50.00–2.26 50.00–2.00 Rwork (%)/Rfree (%)c,d 17.1/19.3 18.3/20.4 Rmsd for bonds (Å) 0.014 0.010 Rmsd for angles (°) 1.507 1.360 Wilson B-factor (Å2) 26.7 23.0 No. of nonhydrogen protein atoms 15,494 15,570 No. of water oxygen atoms 571 732 Ramachandran plot (%) Most favored regions 96.80 96.17 Additional allowed regions 2.63 3.62 Generously allowed regions 0.57 0.21 PDB entry 6JV9 6JVB ↵a Values in parentheses are for the highest resolution shell. About 5% of total reflections are used for highest resolution shell calculations.
↵b Rmerge = Σh Σl |Ihl − 〈Ih〉|/Σh Σl 〈Ih〉, where Ihl is the lth observation of reflection h and 〈Ih〉 is the weighted average intensity for all observations l of reflection h.
↵c Rwork factor = Σh││Fobs(h)│ − │Fcal(h)││/Σh│Fobs(h)│, where Fobs(h) and Fcal(h) are the observed and calculated structure factors for reflection h, respectively.
↵d Rfree factor was calculated same as Rwork factor using the 5% the reflections selected randomly and omitted from refinement.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
