




Article Figures & Data
Figures
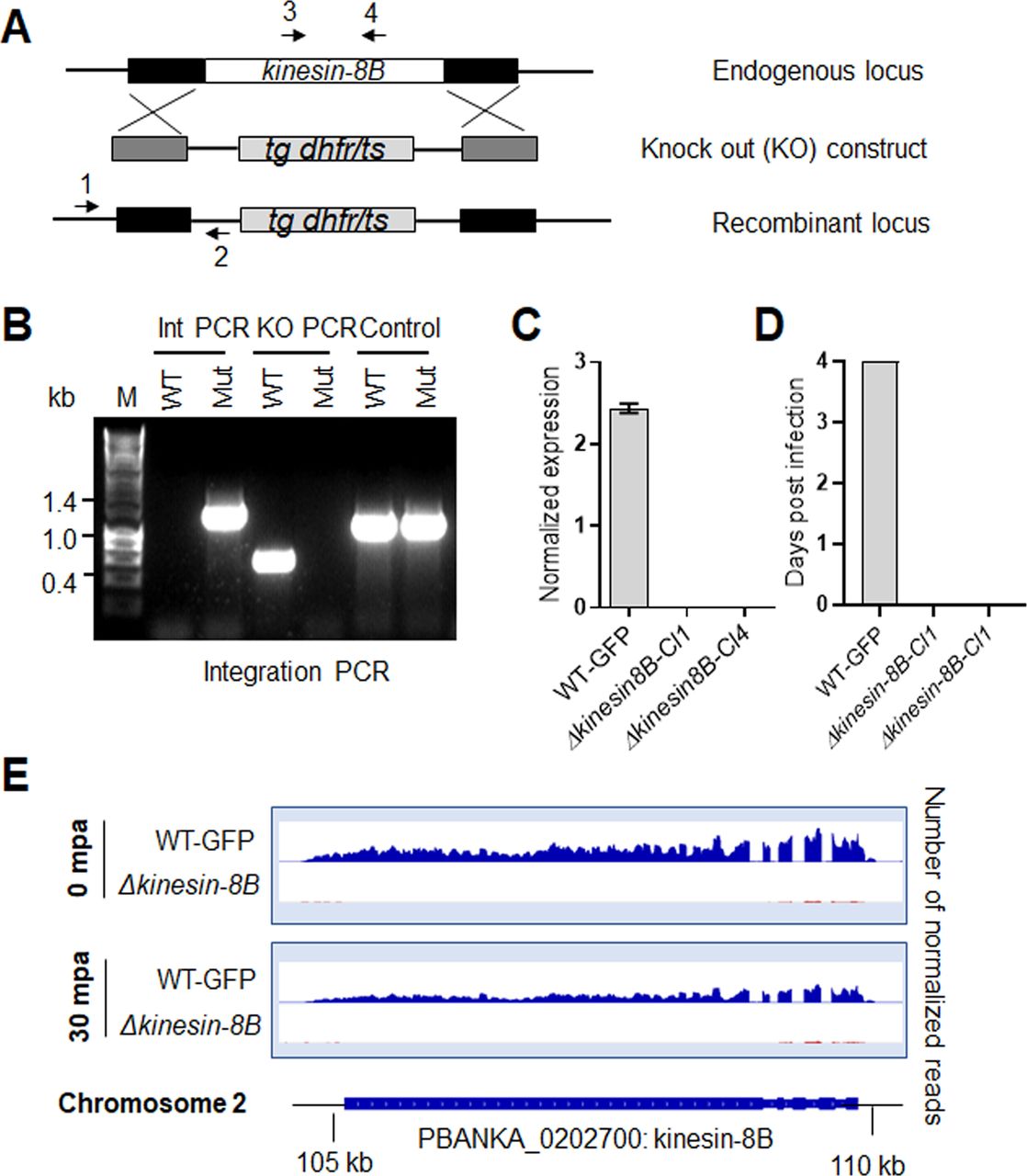
- Figure S1. Generation and genotypic analysis of ∆kinesin-8B parasites.
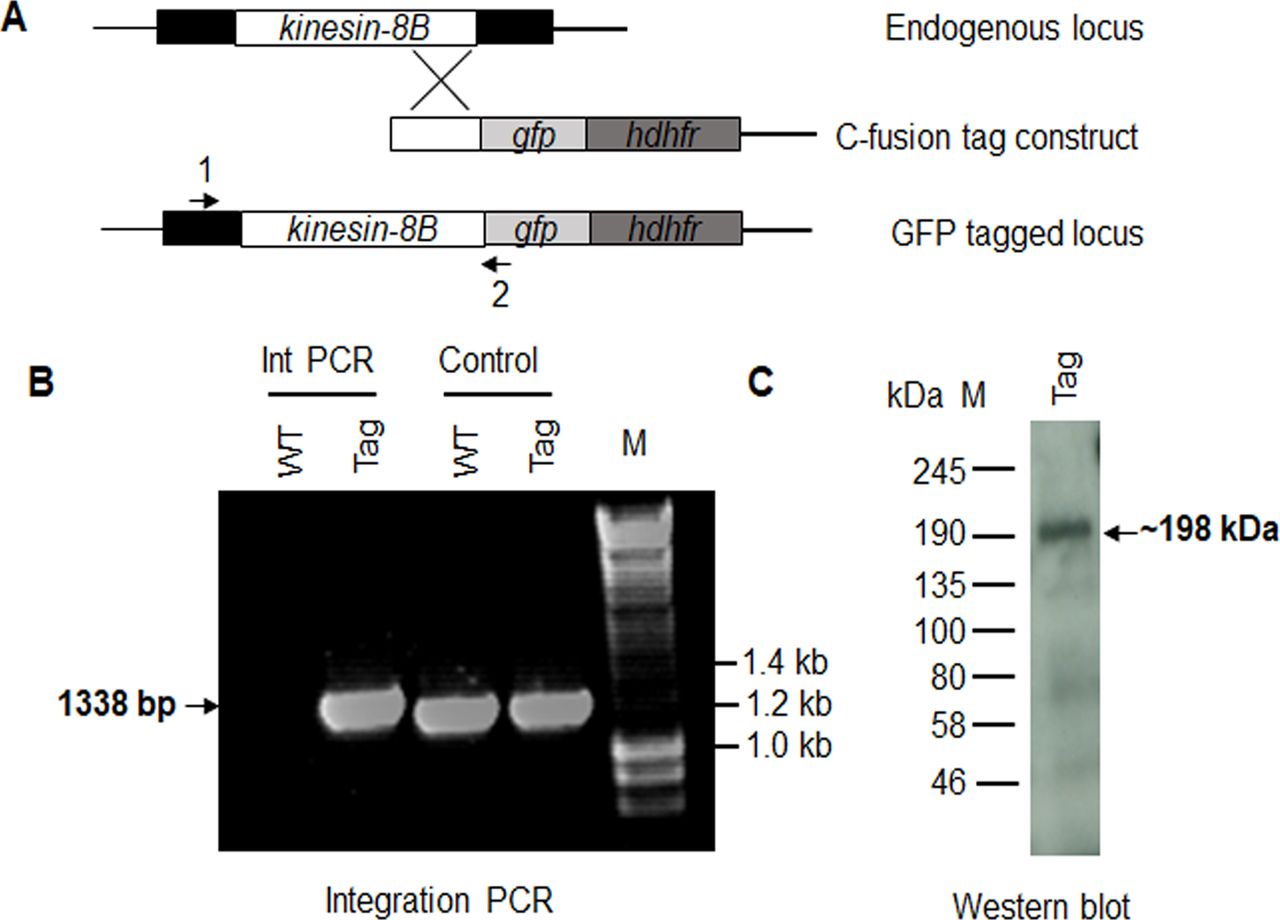
(A) Schematic representation of the endogenous kinesin-8B locus, the targeting knockout construct and the recombined kinesin-8B locus after double homologous crossover recombination. Arrows 1 and 2 indicate PCR primers used to confirm successful integration in the kinesin-8B locus following recombination and arrows 3 and 4 indicate PCR primers used to show deletion of the kinesin-8B gene. (B) Integration PCR of the kinesin-8B locus in WT-GFP and ∆kinesin-8B (Mut) parasites using primers INT N105 and ol248. Integration of the targeting construct gives a band of 1.5 kb. (C) qRT-PCR analysis of transcript in WT-GFP and ∆kinesin-8B parasites. (D) Bite back experiments showing no transmission of Δkinesin-8B parasites, whereas WT-GFP parasites show successful transmission from mosquito to mice. (E) RNAseq analysis confirming lack of transcripts from locus on chromosome 2 in Δkinesin-8B parasites.
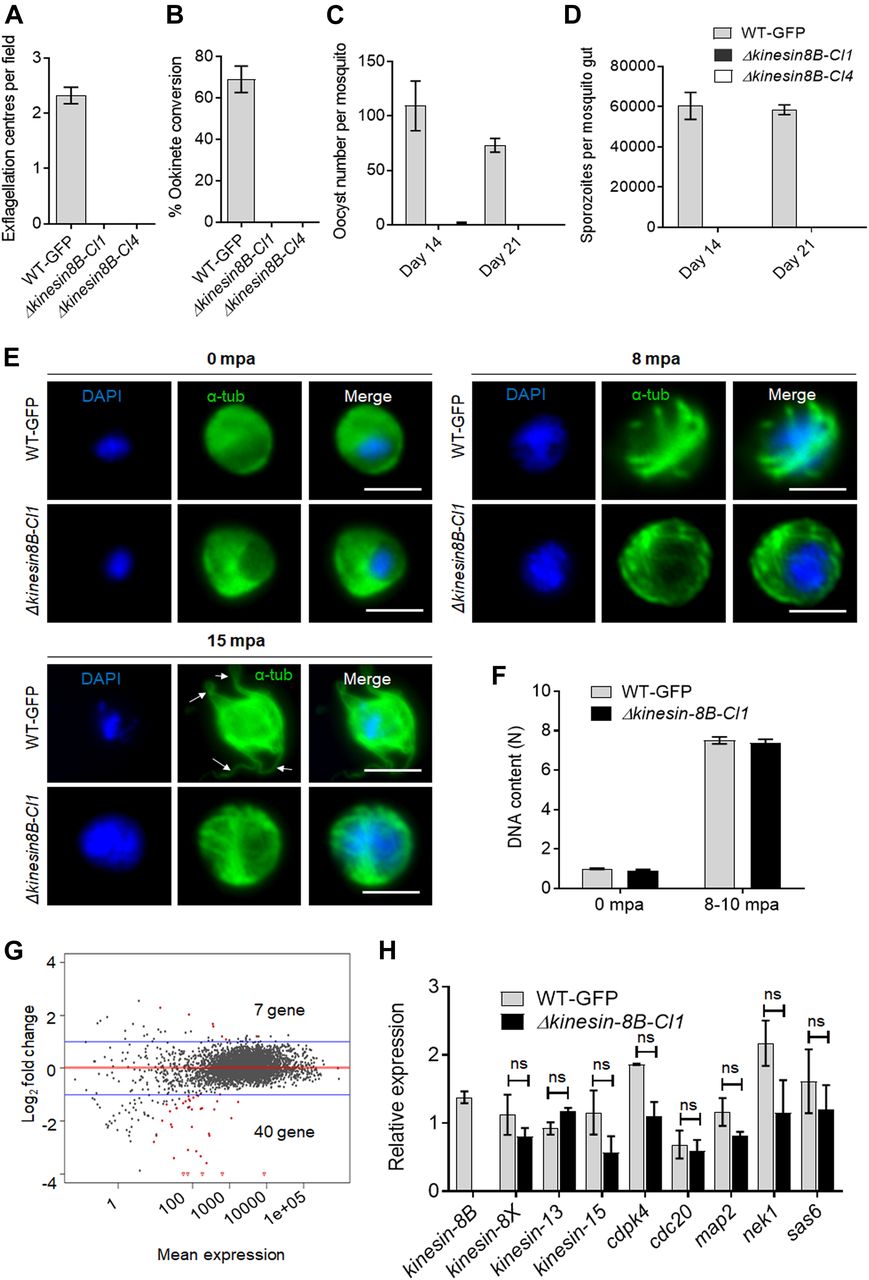
- Figure 1. Kinesin-8B is required for male gamete formation and parasite transmission.
(A) Male gametes (determined by quantifying exflagellation centres) are absent in Δkinesin-8B clones (Cl1 and Cl4) but not in wild-type parasites. Data are means from 15 random independent fields with a 40x objective and three independent replicates. Error bars: ± SEM. (B) Ookinete conversion from zygotes in wild-type and Δkinesin-8B parasites (Cl1 and Cl4). n ≥ 3 independent experiments (>70 cells per experiment for wild type). Error bars: ± SEM. (C) Total number of oocysts of Δkinesin-8B (Cl1 and Cl4) and WT-GFP parasites in mosquito midguts at 14 and 21 dpi. Data are shown as Mean ± SEM, n = 3 independent experiments. (D) Total number of sporozoites in oocysts of Δkinesin-8B (Cl1 and Cl4) and WT-GFP parasites at 14 and 21 dpi. Mean ± SEM. n = 3 independent experiments. (E) In the absence of kinesin-8B, α-tubulin is not reorganised during gametogenesis. IFA images of α-tubulin in WT-GFP and Δkinesin-8B male gametocytes either before activation (0 mpa) or 8 and 15 mpa. DAPI (DNA) is blue. White arrows indicate MT protrusions that occur as the flagellum develops in WT parasites, Scale bar = 5 μm. (F) Fluorometric analyses of DNA content (N) after DAPI nuclear staining. Male gametocytes were at 0 min (non-activated), or 8–10 mpa. The mean DNA content (and SEM) of 10 nuclei per sample are shown. Values are expressed relative to the average fluorescence intensity of 10 haploid ring-stage parasites from the same slide. (G) Differentially expressed genes in Δkinesin-8B parasites compared with WT-GFP parasites. Samples were collected at 0 mpa. (H) qRT-PCR analysis of changes in transcription of selected genes affected in Δkinesin-8B parasites. Error bars: ± SEM. The selected genes have an established or probable role in male gamete development. ns, non significant.
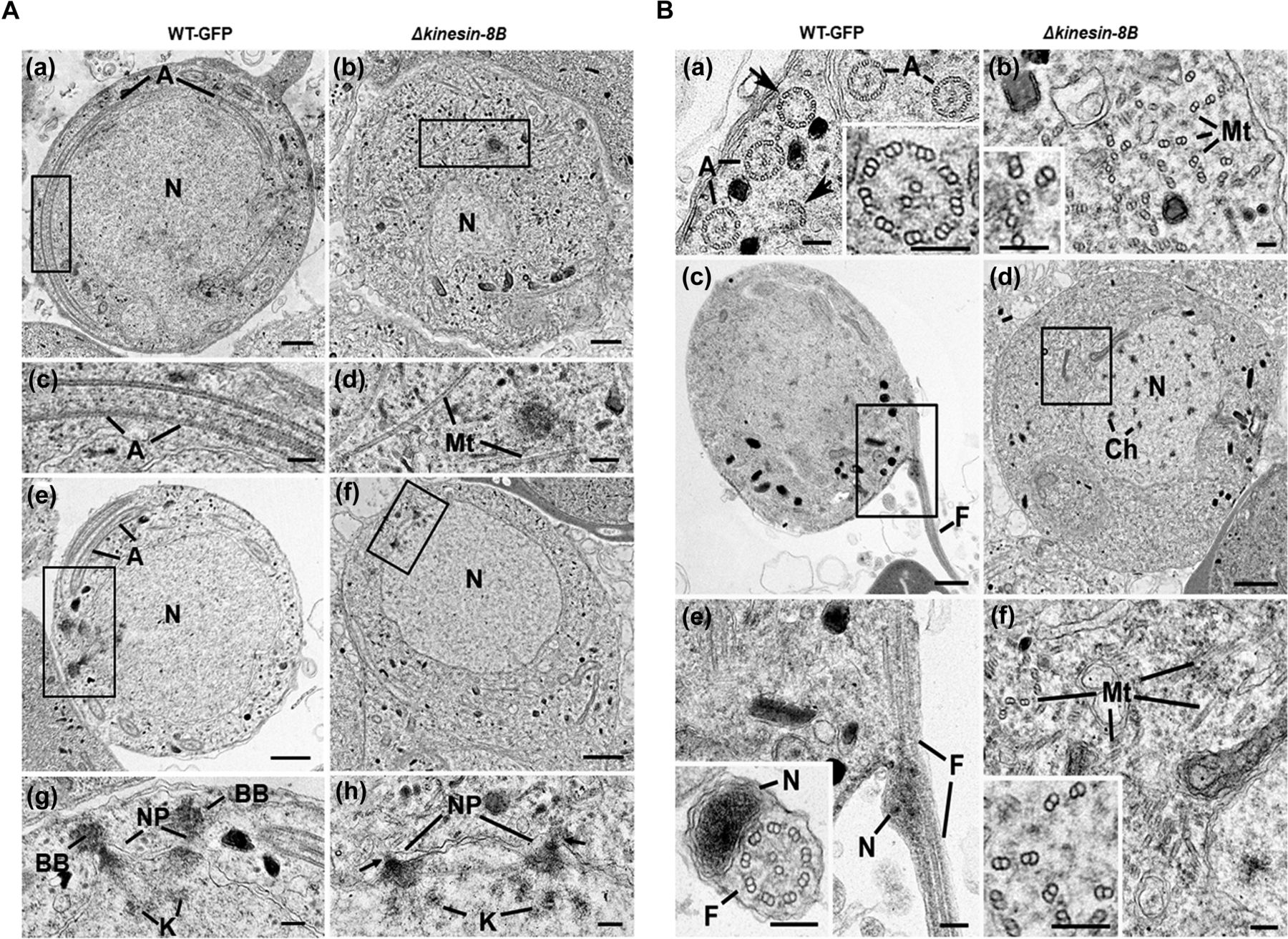
- Figure 2. Ultrastructure analysis reveals defects in basal body and axoneme formation.
(A) Electron micrographs of mid-stage (8 mpa) male gametocytes of WT-GFP (a, c, e, and g) and Δkinesin-8B (b, d, f, and h) parasites. Bars represent 1 μm in (a, b, e, and f) and 100 nm in (c, d, g, and h). (a) Low power image of a WT-GFP male gametocyte showing the large nucleus (N) with cytoplasm containing long axonemes (A) running around the periphery. (b) Low power image of a Δkinesin-8B male gametocyte showing the nucleus (N) and the cytoplasm containing randomly orientated microtubules (Mt). (c) Detail of the enclosed area in panel (a) showing the parallel organization of the microtubules forming an axoneme (A). (d) Detail of the enclosed area in panel B showing randomly orientated microtubules (Mt). (e) WT-GFP male gametocyte showing the large nucleus (N) with multiple nuclear poles, spindle and basal bodies with cytoplasm containing a number of axonemes (A). (f) Δkinesin-8B male gametocyte showing the nucleus with nuclear poles and spindle but no basal bodies. The cytoplasm lacks axonemes but has numerous microtubules (Mt). (g) Detail from the enclosed area in (e) showing basal bodies (BB), the nuclear pole (NP) connected by spindle microtubules with attached kinetochores (K). (h) Detail from the enclosed area in (f) showing the nuclear poles (NPs) connected by spindle microtubules with attached kinetochores (K). Note the absence of basal bodies associated with the nuclear poles (arrows). (B) Electron micrographs of mid (8 mpa) and late (15 mpa) male gametocytes of WT-GFP (a, c and e) and mutant (b, d and f). Bar represent 1 μm in (c and d) and 100 nm in all other micrographs. (a) Part of the peripheral cytoplasm of a mid-stage (8 mpa) WT-GFP gametocyte showing a cross section through a number 9+2 axonemes (A). Note the presence of incomplete axonemes (arrows). Inset. Detail of a cross sectioned axoneme showing the 9+2 organisation of the microtubules. (b) Part of the cytoplasm of a mid-stage mutant male gametocyte showing numerous randomly orientated microtubules (Mt). Inset. Detail showing the presence of both doublet and single microtubules. (c) Late stage (15 mpa) male gametocyte showing the partial formation of a male gamete by exflagellation. F, Flagellum. (d) Late mutant male gametocyte showing the early chromatin condensation and the absence of axonemes in the cytoplasm. (e) Detail of the enclose area in (c) showing the flagellum (F) and associated nucleus (N) protruding from the surface of the male gametocyte. Inset. Cross-section through a free male gamete showing the electron-dense nucleus (N) and the classical 9+2 flagellum. (f) Detail of the enclosed area in (d) showing the numerous randomly orientated microtubules (Mt). Inset. Detail showing the disorganisation of the doublet microtubules.
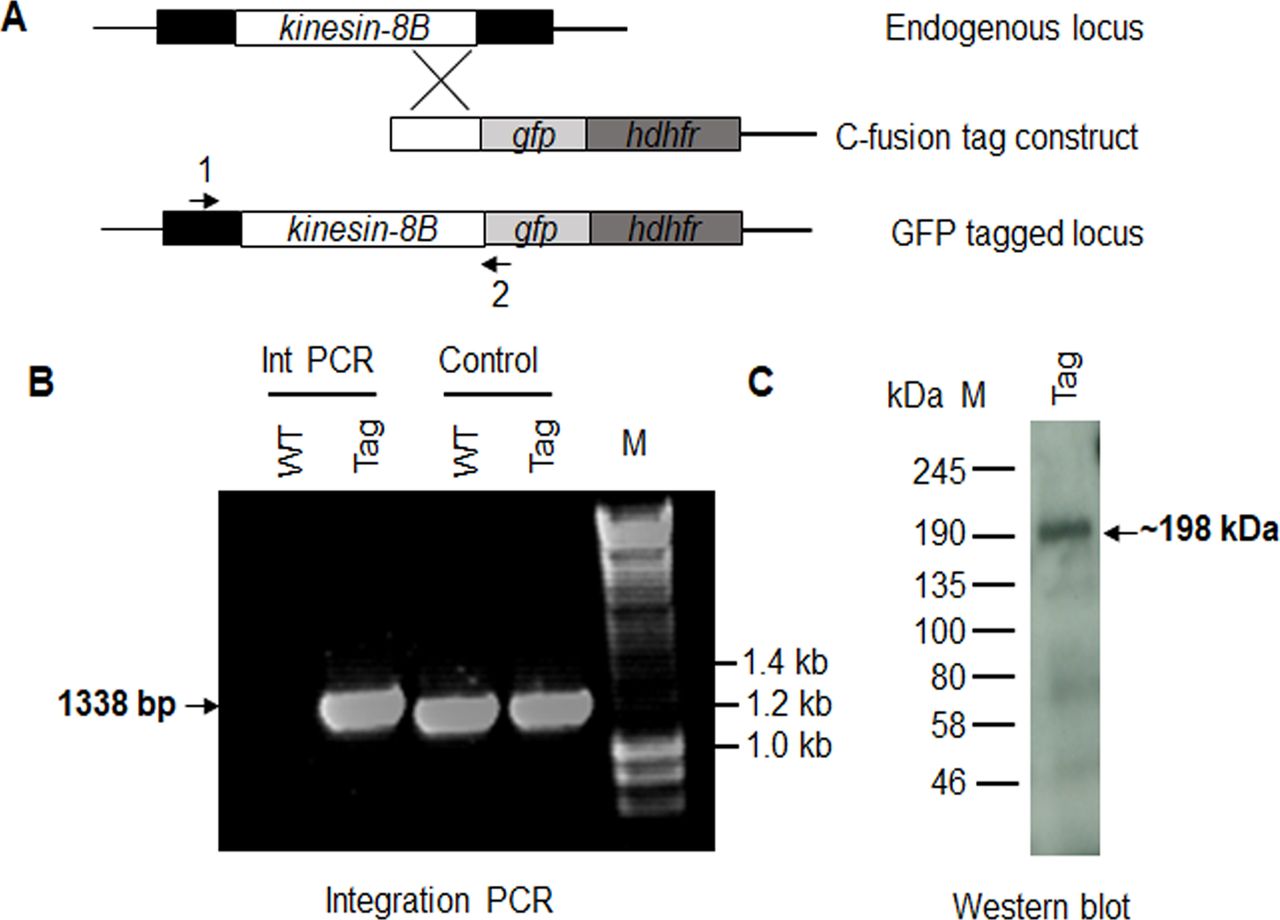
- Figure S2. Generation and genotypic analysis of kinesin-8B–GFP.
(A) Schematic representation of the endogenous pbkinesin-8B locus, the GFP-tagging construct, and the recombined kinesin-8B locus after single homologous recombination. Arrows 1 and 2 indicate the position of PCR primers used to confirm successful integration of the construct. (B) Diagnostic PCR of kinesin-8B and WT parasites using primers IntT199 (arrow 1) and ol492 (arrow 2). Integration of the kinesin-8B tagging construct gives a band of 1,338 bp. Tag = kinesin-8B–GFP parasite line. (C) Western blot of kinesin-8B–GFP (∼198 kD) protein illustrates the presence of the protein in gametocyte stage.
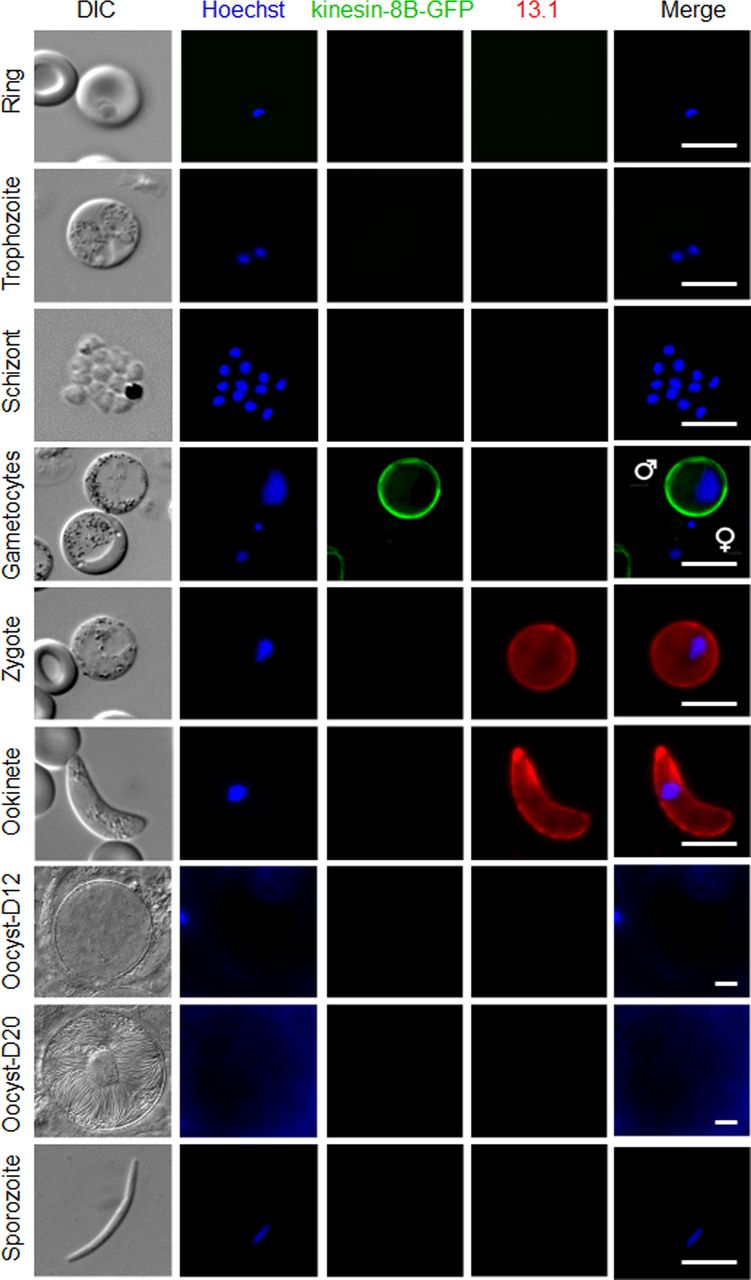
- Figure S3. Expression and localisation of kinesin-8B–GFP throughout the life cycle of P. berghei.
Expression of kinesin-8B–GFP during blood schizogony (ring, trophozoite, and schizonts), male and female gametocytes (8 mpa), zygote, ookinete, oocysts (day 12 and day 20), and sporozoites. Gametocytes shown here are at 8 mpa. 13.1, a cy3-conjugated antibody which recognises P28 on the surface of activated females, zygotes, and ookinetes, was used with the sexual stages. Scale bar = 5 μm.
Source data are available for this figure.
Source Data for Figure S3[LSA-2019-00488_SdataFS3_1.pdf][LSA-2019-00488_SdataFS3_2.pptx]
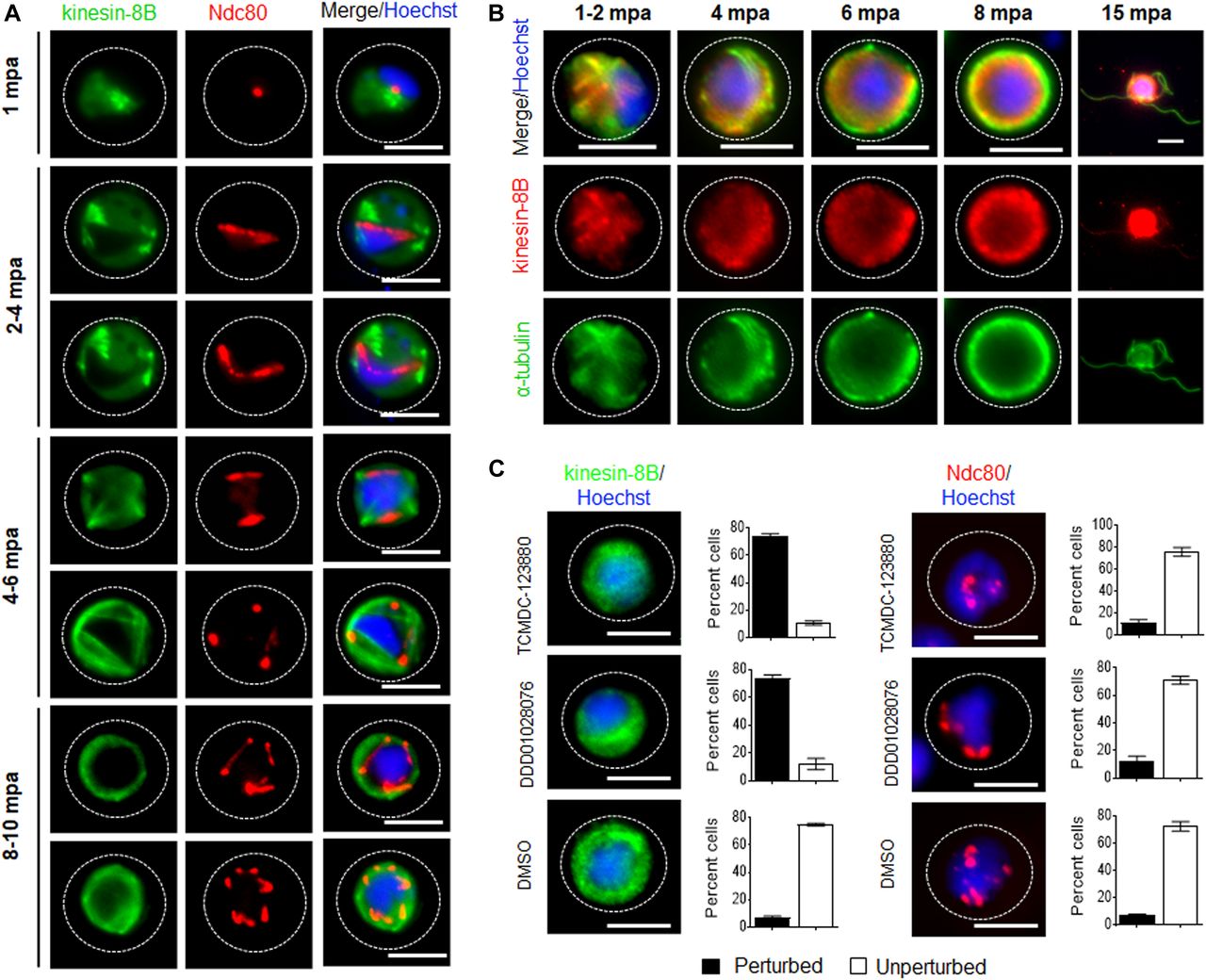
- Figure 3. The location of kinesin-8B in relation to that of the kinetochore (Ndc80) and microtubule (α-tubulin) markers.
(A) The location of kinesin-8B–GFP (green) in relation to the kinetochore marker, Ndc80-mCherry (red) during male gamete formation. The cytoplasmic location of kinesin-8B contrasts with the nuclear location of Ndc80 during chromosome replication and segregation, indicating that kinesin-8B is not associated with the mitotic spindle. The non-clonal lines expressing kinesin-8B–GFP and Ndc80-mCherry were crossed and then individual parasites expressing both markers were analysed by live fluorescence microscopy. (B) Indirect IFA showing co-localisation of kinesin-8B (red) and α-tubulin (green) in male gametocytes at 1 to 2, 4, 6, 8, and 15 mpa. (C) Antimalarial molecules block the dynamic distribution of kinesin-8B showing the resulting phenotype of compound addition at 4 mpa. In contrast to the effect on kinesin-8B–GFP distribution, no significant effect was seen on Ndc80–RFP. Inhibitors were added at 4 mpa and parasites were fixed at 8 mpa. Scale bar = 5 μm.
- Figure 4. The dynamic location of kinesin-8B in basal body and axoneme formation.
(A) Cytoplasmic location of kinesin-8B–GFP (green) by live cell imaging in real time during male gamete development. DNA is stained with Hoechst 33,342 (blue). Before activation (0 min), kinesin-8B has a diffused location in male gametocyte cytoplasm. 1–2 mpa, it accumulates at one end of the nucleus and forms four foci reminiscent of the “tetrad of basal body” defined by electron microscopy (Sinden et al, 1976). These tetrad foci are duplicated within 2–4 mpa, and by 4–6 mpa fibre-like structures, representing axonemal growth decorated with kinesin-8B–GFP, extend from these tetrads to make a basket-like structure around the nucleus, which is completed by 8–10 mpa. After exflagellation, kinesin-8B–GFP is located along the length of the flagellum in the free male gamete (15 mpa, inset). For each time point, a cartoon guide is presented. (B) Still images (at every 5 s) of tetrad-foci duplication and the start of axonemal growth decorated with kinesin-8B–GFP at 2–3 mpa (Fig 4B1; Video 1) and 3–4 mpa (Fig 4B2; Video 2). Scale bar = 5 μm.
- Figure 5. Schematic model of kinesin-8B location on basal bodies and axonemes assembly during the male gamete development.
Kinesin-8B accumulates at one end of the nucleus within 1 mpa of male gametocytes, probably near the MT organizing centre (MTOC) and develops into four foci corresponding to the tetrad of basal bodies (BBs, green). These tetrad foci of kinesin-8B duplicate and separate from each other within 2–4 mpa. Each of these foci is a basal body that serves as a platform for axonemal MT assembly. The fibre-like structure decorated with kinesin-8B (green) emerges from these foci, showing their association with axoneme assembly. Meanwhile, DNA replication accompanies basal body duplication, with associated mitotic spindles (yellow) and kinetochores (red) via the spindle pole (Sinden et al, 1976). The axonemal fibre-like structures develop further into a basket-like structure around the nucleus by 6–10 mpa. Axoneme formation and DNA replication are completed by this time (8–10 mpa) and a haploid genome connected with a basal body, is pulled into an emerging gamete.





Supplementary Materials
- Video 1
Time-lapse video showing duplicated tetrad foci of kinesin-8B moving apart from each other in 50 s.Download video
- Video 2
Dynamic movement of duplicated tetrad foci and emerging fibre-like structure of kinesin-8B in 50 s.Download video
Table S3 Oligonucleotides used in this study.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Related Articles
Cited By...
- Microtubule inner proteins of Plasmodium are essential for transmission of malaria parasites
- A multistage Plasmodium CRL4WIG1 ubiquitin ligase is critical for the formation of functional microtubule organisation centres in microgametocytes
- Atypical flagella assembly and haploid genome coiling during male gamete formation in Plasmodium
- Rita Tewari: malaria parasite cell division
- Plasmodium SAS4: basal body component of male cell which is dispensable for parasite transmission
- Mechanochemical tuning of a kinesin motor essential for malaria parasite transmission
- Real-time dynamics of Plasmodium NDC80 reveals unusual modes of chromosome segregation during parasite proliferation
- A divergent cyclin/cyclin-dependent kinase complex controls the atypical replication of a malaria parasite during gametogony and transmission