



Article Figures & Data
Figures
- Figure S1. The basic AJ and TJ structural integrity is maintained in vinculin KO epithelial cell sheets.
(A) Western blots for AJ and TJ proteins (vinculin, E-cadherin, ZO-1, and claudin-4) and β-actin in WT, vinculin KO, and REV Eph4 epithelial cells. Representative images from three independent experiments are shown. (B) Paracellular barrier function for ions, shown by the TER of WT and various clones of vinculin KO Eph4 epithelial cell sheets. Results from three independent experiments are shown as the mean ± SEM (n = 3/group). (C) Freeze-fracture replica electron microscopic images of TJ strands in WT and vinculin KO Eph4 epithelial cell sheets. Representative images from two independent experiments are shown. Scale bars: 100 nm. (D) Quantification of junctional depth. TJ strands were transected by lines perpendicular to strands. Junctional depths were measured from the most luminal strand to the most distal strand. Images were recorded from three independent experiments and analyzed. There is no significant difference between junctional depth of WT and vinculin KO Eph4 cells. (E) Quantification of strand number. Strand number was measured from the most luminal strand to the most distal strand. Images were recorded from three independent experiments and analyzed. There is no significant difference between junctional depth of WT and vinculin KO Eph4 cells.
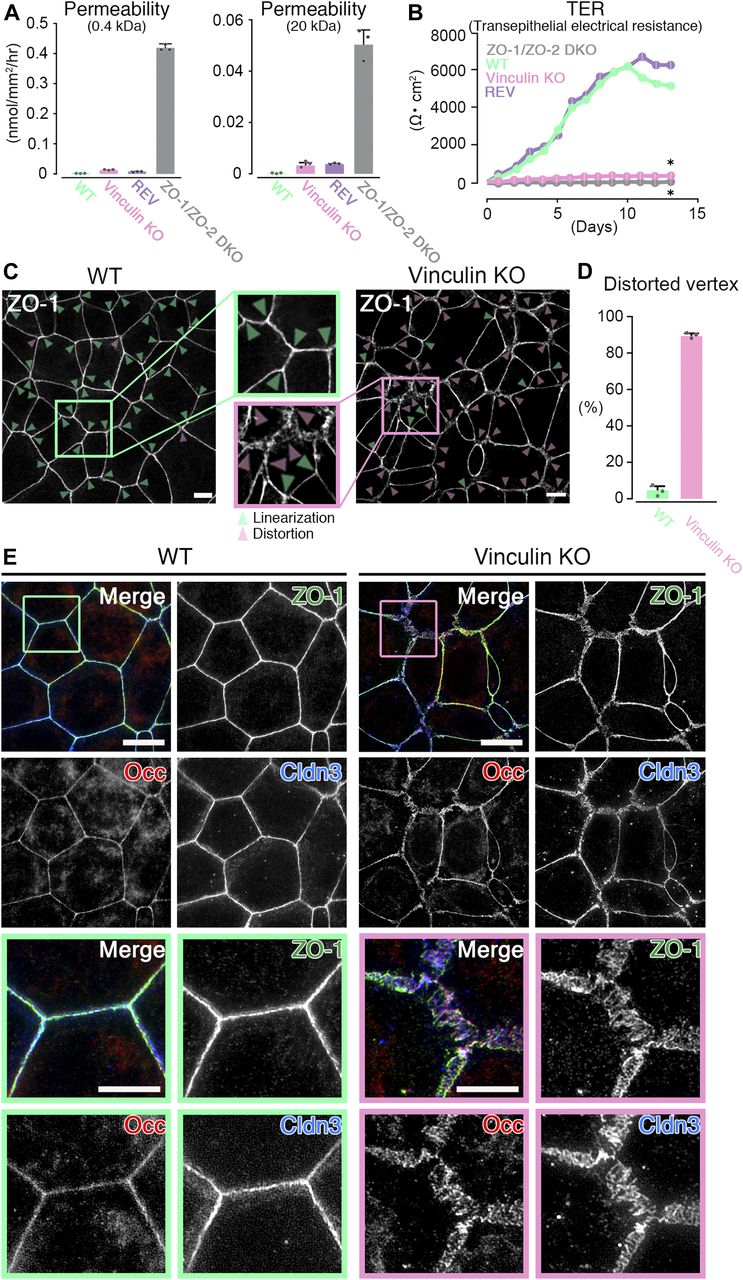
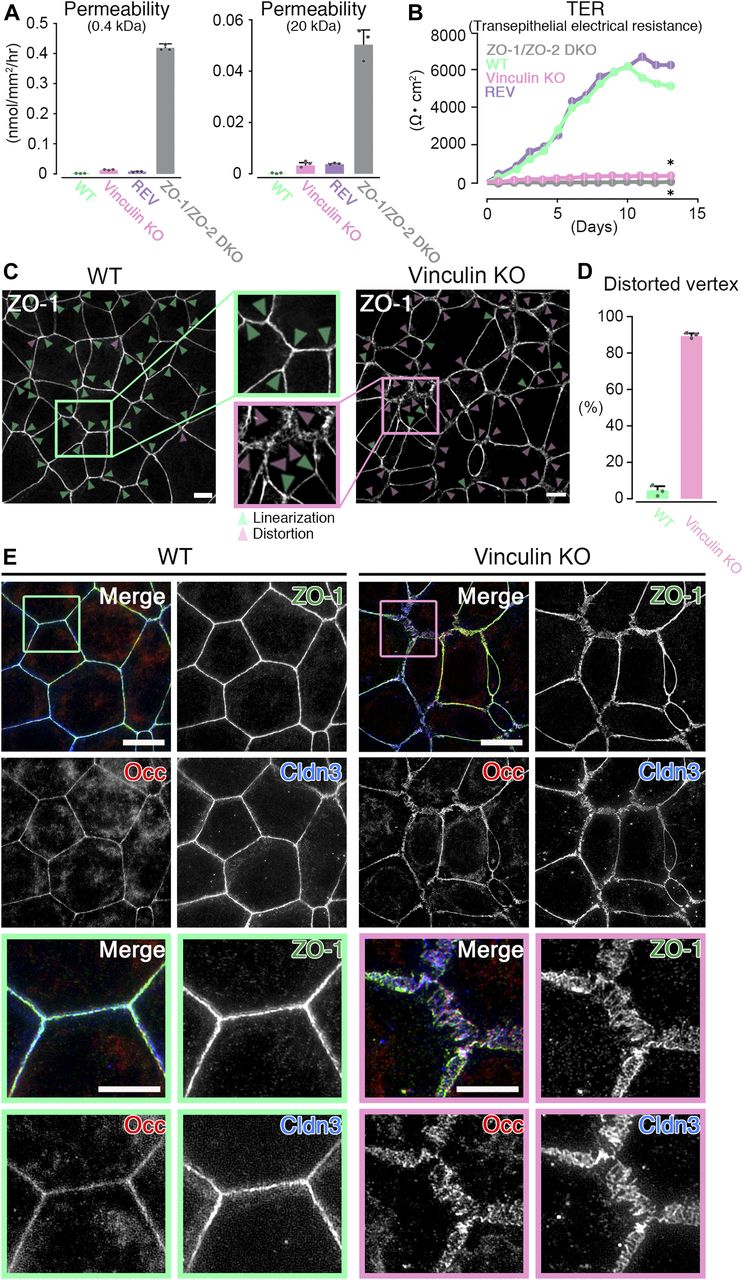
- Figure 1. The paracellular barrier function for large solutes is largely maintained, but that for ions is lost in vinculin KO epithelial cell sheets.
(A) Paracellular barrier function for large solutes, shown by paracellular tracer flux assays using 0.4-kD fluorescein (left) and 20-kD FD-20 (right), in WT, vinculin KO, REV, and ZO-1/ZO-2 DKO Eph4 epithelial cell sheets. Under the condition of full confluency (14 d of culture on a filter), the paracellular barrier function for fluorescein and FD-20 was largely maintained in vinculin KO cell sheets compared with WT and REV epithelial cell sheets. In contrast, in ZO-1/ZO-2 DKO cell sheets, which lacked TJs, the paracellular barrier function for fluorescein and FD-20 was severely defective. Results from three independent experiments are shown as the mean ± SEM (n = 3/group). (B) Paracellular barrier function for ions, shown by the TER of WT, vinculin KO, REV, and ZO-1/ZO-2 DKO Eph4 epithelial cell sheets. Under the condition of full confluency (13 d of culture on a filter), in the WT and REV cell sheets, the paracellular barrier function for ions was very high (TER of 5,500 ∼ 6,500 Ω·cm2). In contrast, in the vinculin KO cell sheets, the paracellular barrier function for ions was almost completely lost, with a TER (500–600 Ω·cm2) almost the same as that of the TJ-less ZO-1/ZO-2 DKO epithelial cell sheets. Results from three independent experiments are shown as the mean ± SEM (n = 3/group). P-values were calculated using a two-tailed independent t test, and P < 0.05 was considered significant. *P < 0.05. (C) Confocal super-resolution immunofluorescence images for ZO-1, a TJ protein, in WT (left) and vinculin KO (right) Eph4 epithelial cell sheets. In WT cell sheets, the ZO-1 signals showed a continuous, belt-like linear junctional pattern (green arrowheads: linear ZO-1 distribution). In contrast, in vinculin KO epithelial cell sheets, the continuous linear junctional pattern of ZO-1 was stochastically distorted to become expanded in the tricellular point (magenta arrowheads: distorted ZO-1 distribution). Scale bars: 10 μm. (D) The ratio of distorted vertices to all the tricellular vertices of each cell sheets was calculated by representative images from three independent experiments and described. (E) Confocal super-resolution immunofluorescence images for TJ proteins ZO-1, occludin (Occ), and claudin-3 (Cldn3) in WT (left) and vinculin KO (right) Eph4 epithelial cell sheets. In WT epithelial cell sheets, ZO-1 (green), occludin (red), and claudin-3 (blue) showed a continuous belt-like linear junctional pattern of immunofluorescence. In contrast, in vinculin KO epithelial cell sheets, the continuous linear junctional patterns of ZO-1 (green), occludin (red), and claudin-3 (blue) were stochastically distorted to then become expanded. Lower images are magnifications of the boxed regions in the upper images. Representative images from three independent experiments are shown. Scale bars: 10 μm.
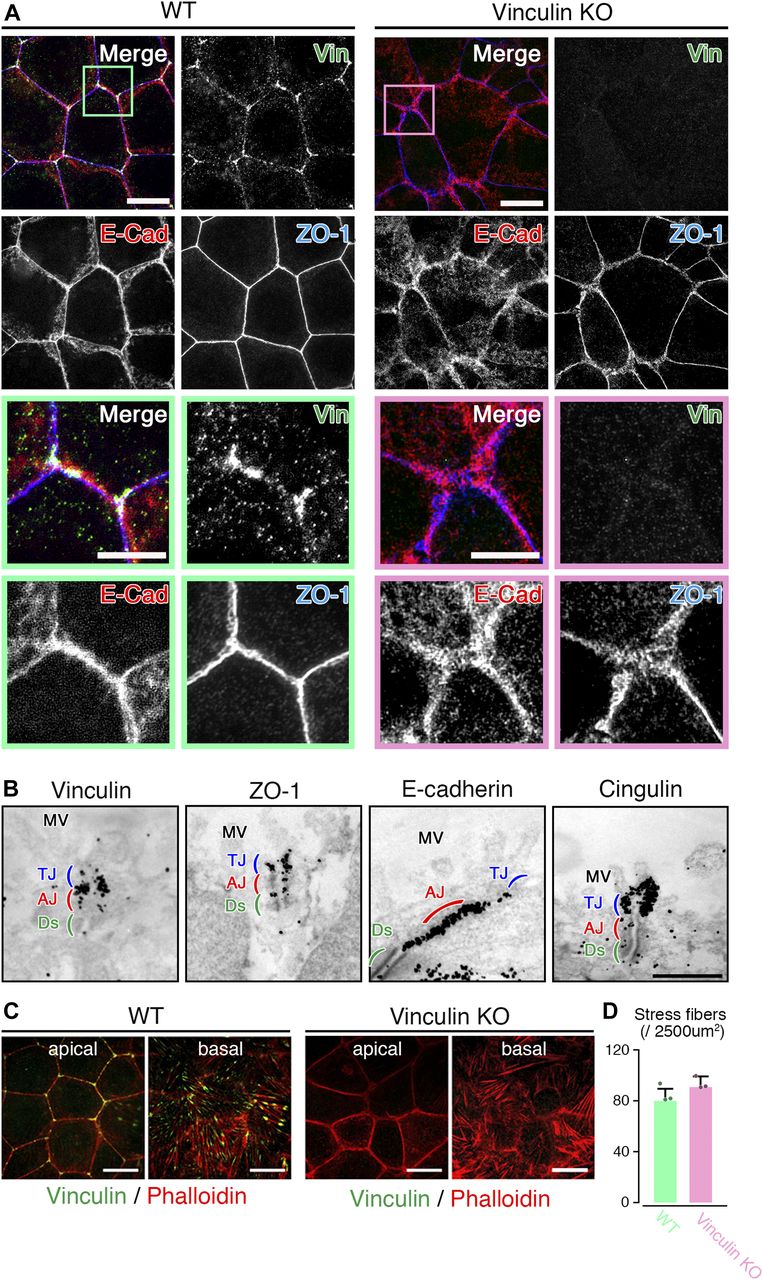
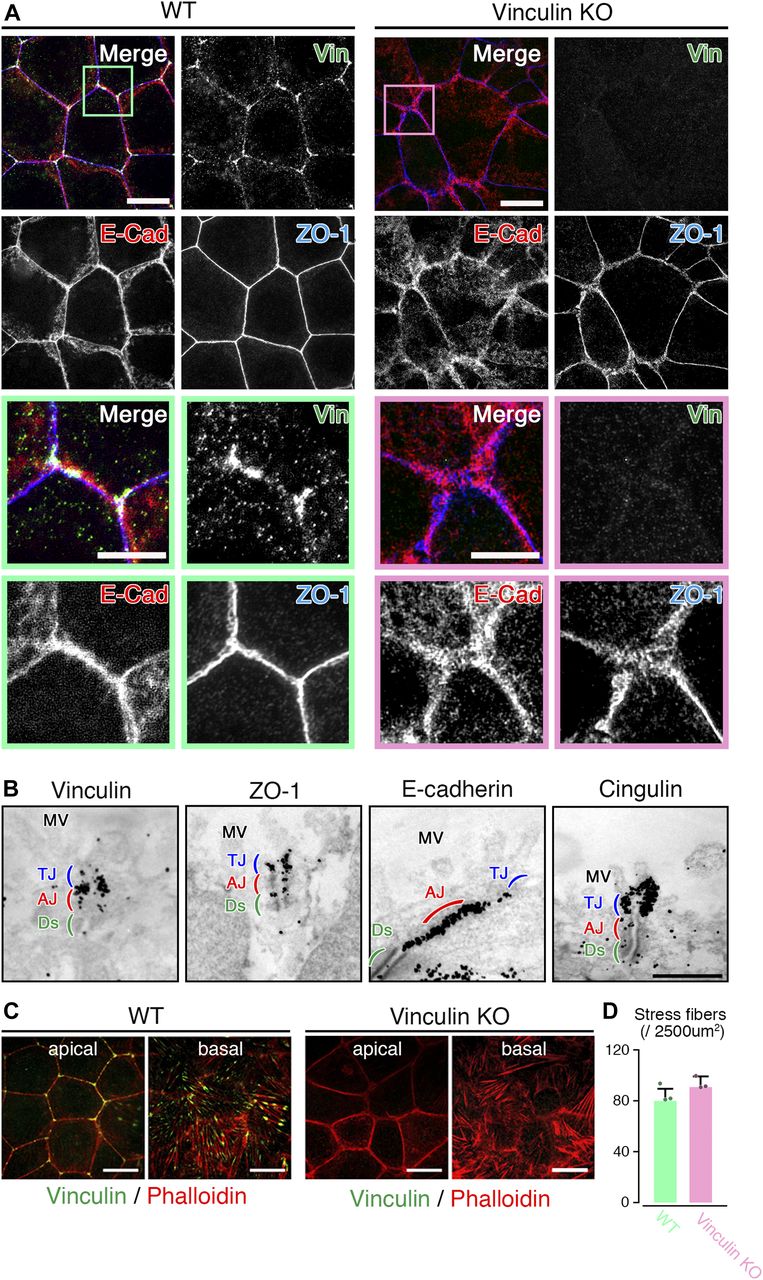
- Figure 2. The linear distribution patterns of AJ and TJ proteins are stochastically distorted to become expanded in vinculin KO epithelial cell sheets.
(A) Confocal super-resolution immunofluorescence images of an AJ protein E-cadherin (E-Cad), a TJ protein ZO-1, and vinculin (Vin) in WT (left) and vinculin KO (right) Eph4 epithelial cell sheets. In WT epithelial cell sheets, vinculin (green) was co-localized with E-cadherin (red) and ZO-1 (blue) at the most apical parts of the lateral membranes and was particularly concentrated at tricellular junctions. In vinculin KO epithelial cell sheets, the vinculin was completely lost and the distribution pattern of E-cadherin (red) was stochastically distorted to then become expanded. Lower images are magnified images of the boxed regions in the upper images. Representative images from three independent experiments are shown. Scale bars: 10 μm. (B) Immunoelectron microscopy images of AJ and TJ proteins vinculin, ZO-1, E-cadherin, and cingulin in WT Eph4 epithelial cell sheets. Immunogold signals for vinculin were predominantly observed in the AJ (red) with a very few signals in the TJ (blue). The AJ protein E-cadherin and TJ protein cingulin were predominantly localized to the AJ and TJ, respectively. Notably, immunogold signals for ZO-1 were observed in both the TJ and the AJ. Representative images from three independent experiments are shown. MV, microvilli; Ds, desmosome. Scale bar: 500 nm. (C) Confocal super-resolution immunofluorescence images of phalloidin, an F-actin marker, and vinculin in WT (left) and vinculin KO (right) Eph4 epithelial cell sheets. In WT epithelial cell sheets, vinculin (green) accumulated especially at tricellular junctions in the apical side of the cells and was also localized to the edge of stress fibers, indicated by phalloidin (red), at the basal side of the cells. Scale bars: 10 μm. (D) The number of stress fiber in the z projection image of the basal side of the cells recorded by super-resolution confocal microscopy was measured. Representative images from three independent experiments were used. There was no significant difference in the stress fiber formation between WT and vinculin KO epithelial cell sheets.
- Figure S2. Vinculin at the AJ is connected to the TJ via ZO-1 in WT epithelial cell sheets.
(A) Immunoelectron microscopy images of an AJ protein β-catenin and a TJ protein occludin in WT Eph4 epithelial cell sheets. Immunogold signals for occludin were predominantly observed in TJs (blue) and those for β-catenin were mostly observed in AJs (red). Representative images from three independent experiments are shown. MV, microvilli; Ds, desmosome. Scale bars: 500 nm. (B) Confocal super-resolution immunofluorescence images of AJ and TJ proteins vinculin (VIN), ZO-1, and E-cadherin (E-Cad) in REV Eph4 epithelial cell sheets. Linear distribution patterns of vinculin (green), E-cadherin (red), and ZO-1 (blue) along cell–cell junctions were observed in the REV Eph4 cell sheets. Lower images are magnified images of the boxed regions in the upper image. Representative images from three independent experiments are shown. Scale bars: 10 μm. (C) Co-immunoprecipitation of vinculin with ZO-1. GFP-tagged ZO-1 and HA-tagged full-length vinculin or its domains (G1, G2–G4, or tail) were used to co-transfect HEK293 cells and their extracts were subjected to pull-down assays with an anti-HA or anti-GFP antibody. GFP-tagged ZO-1 interacted with the HA-tagged G2–G4 domain of vinculin and full-length vinculin. Representative images from three independent experiments are shown. (D) Immunoelectron microscopic images of a TJ protein ZO-1 in vinculin KO Eph4 epithelial cell sheets. Scale bars: 500 nm.
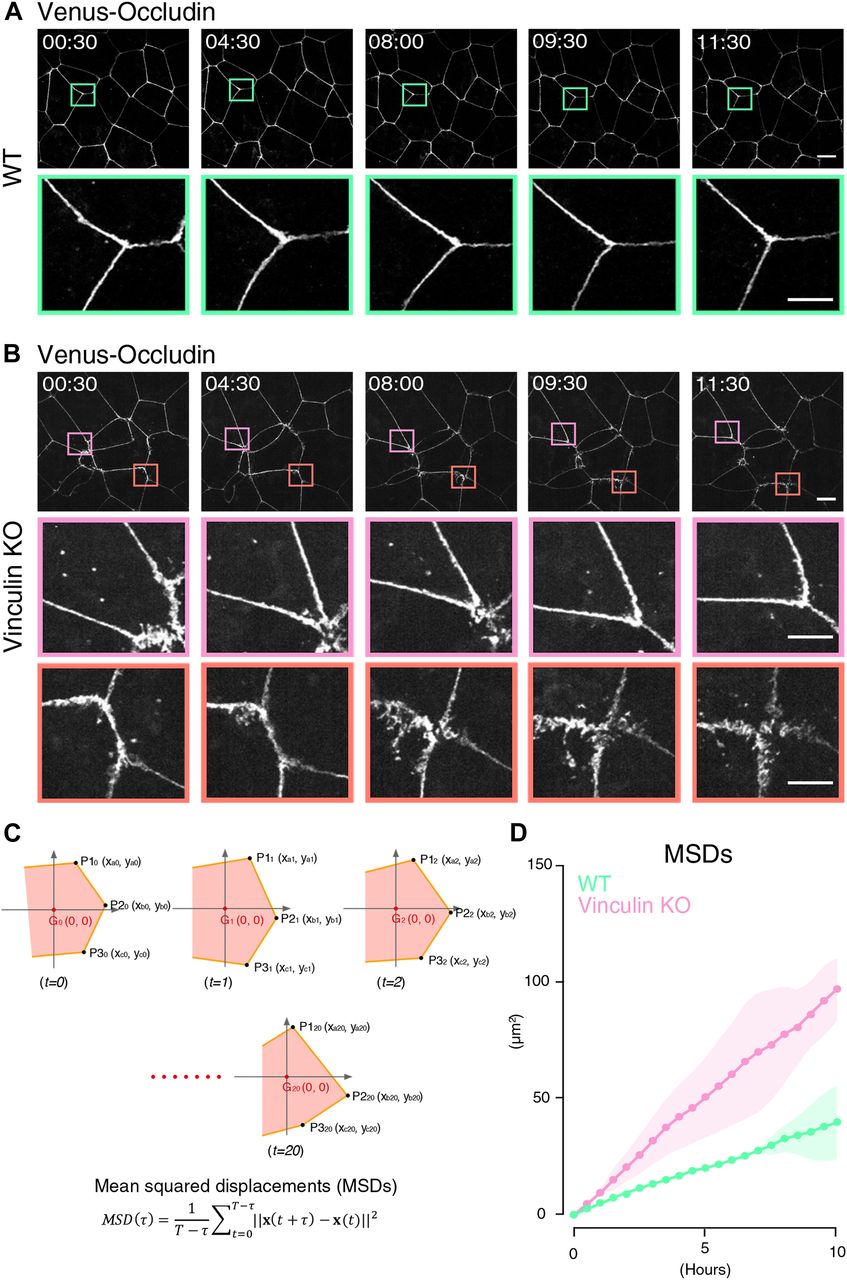
- Figure 3. The transition between linear and distorted TJ patterns in vinculin KO epithelial cell sheets is dynamic.
(A) Live cell imaging of WT Eph4 epithelial cells stably expressing Venus-tagged occludin as a marker for TJs (see Video 1). In Venus–occludin-expressing WT epithelial cell sheets, the cell shape and continuous linear distribution pattern of Venus–occludin were maintained. Lower images are magnifications of the boxed regions in the upper images. Representative time-lapse images of Venus–occludin at the indicated time points from three independent experiments are shown. Time is shown in hours: minutes. Scale bars: 10 μm. (B) Live cell imaging of vinculin KO Eph4 epithelial cells stably expressing Venus-tagged occludin as a marker for TJs (see Video 2). In Venus–occludin-expressing vinculin KO epithelial cell sheets, the TJ patterns showed much more movement than the Venus–occludin-expressing WT epithelial cells. Notably, stochastic transitions from the distorted to linear pattern of Venus–occludin (magenta boxes) and from the linear to distorted pattern of Venus–occludin (orange boxes) were observed in Venus–occludin-expressing vinculin KO cell sheets. Lower images are magnifications of the boxed regions in the upper images. Representative time-lapse images of Venus–occludin at the indicated time points from three independent experiments are shown. Time is shown in hours: minutes. Scale bars: 10 μm. (C) Calculation of the tMSD of the vertices of Venus–occludin-expressing WT and vinculin KO Eph4 epithelial cells. (D) Quantitative analysis of the tMSD of the vertices of Venus–occludin-expressing WT and vinculin KO Eph4 epithelial cells. We selected epithelial cells with similar areas (Fig S3A). The tMSD of the vertices of Venus–occludin-expressing vinculin KO epithelial cells was larger than that of Venus–occludin-expressing WT epithelial cells (Fig 3C), confirming that the vinculin KO epithelial cells showed much more movement than the WT epithelial cells. Results from three independent experiments are shown as mean ± SEM (n = 3/group).
- Figure S3. Mechanical fluctuation induces the distorted AJ and TJ patterns in vinculin KO epithelial cell sheets.
(A) Quantitative analysis of the changes in areas of WT and vinculin KO Eph4 epithelial cells stably expressing Venus-tagged occludin as a marker for TJs, which were used for analysis of the change of tMSD (Fig 3C and D). (B) Confocal super-resolution immunofluorescence images of AJ and TJ proteins vinculin (green), E-cadherin (red), and ZO-1 (blue) in WT Eph4 epithelial cell sheets treated with blebbistatin, an actomyosin tension-relaxing reagent, or DMSO. Representative images from three independent experiments are shown. Scale bars: 10 μm. (C) Confocal super-resolution immunofluorescence images of AJ and TJ proteins vinculin (green), E-cadherin (red), and ZO-1 (blue) in vinculin KO Eph4 epithelial cell sheets treated with blebbistatin or DMSO. Blebbistatin but not DMSO restored the linear distribution patterns of E-cadherin (red) and ZO-1 (blue) along cell–cell junctions in vinculin KO cell sheets. Representative images from three independent experiments are shown. Scale bars: 10 μm.
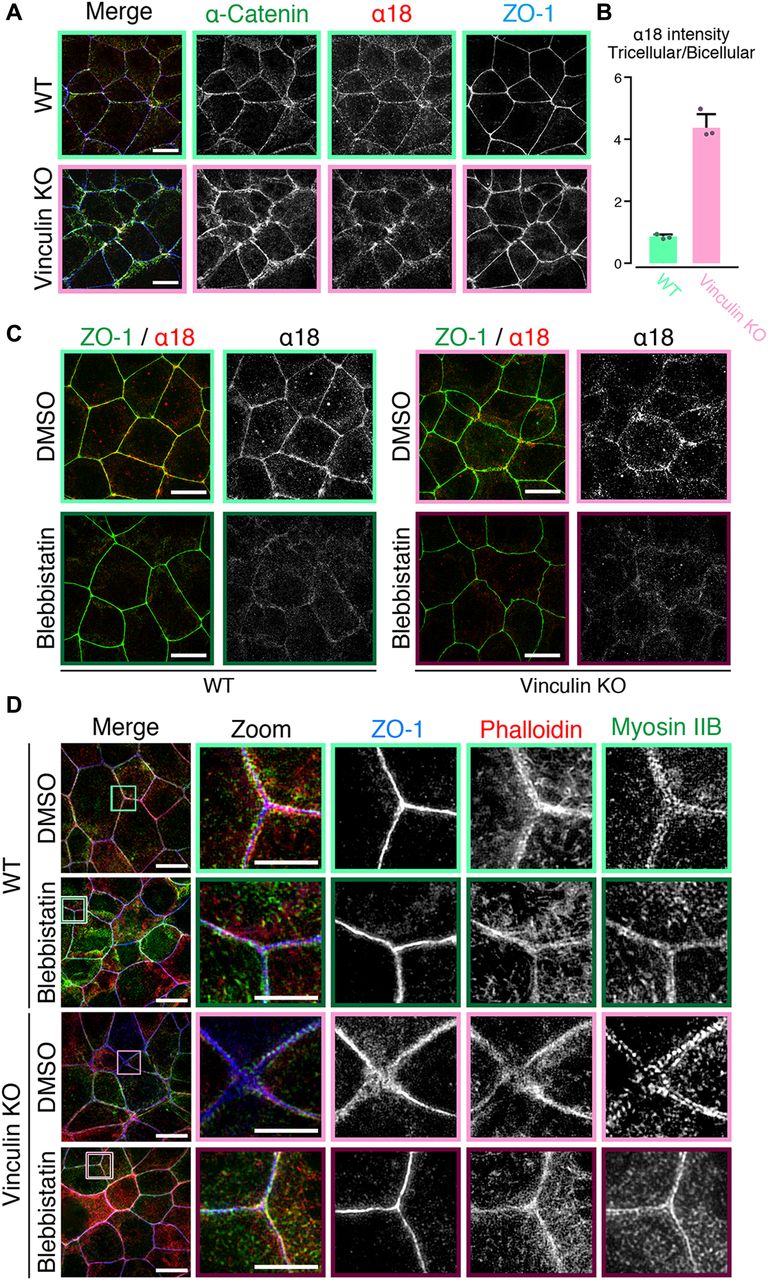
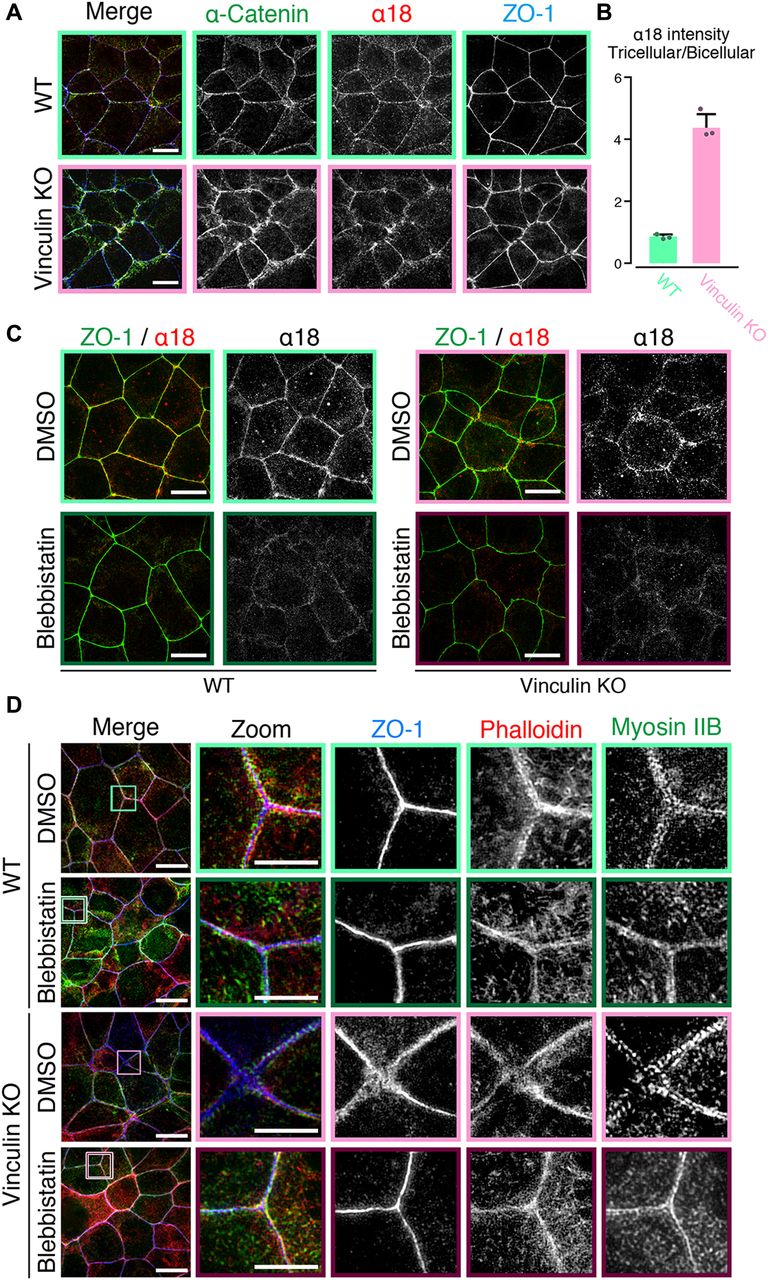
- Figure 4. Mechanical fluctuation induces the distorted TJ patterns in vinculin KO epithelial cell sheets.
(A) Confocal super-resolution immunofluorescence images of α18 antigen, which indicates mechanical fluctuation, in WT (upper) and vinculin KO (lower) Eph4 epithelial cell sheets. The α18-antigen signals were concentrated at tricellular regions, as shown by a TJ protein ZO-1 (red), and DAPI-stained nuclei (blue), in WT and vinculin KO epithelial cell sheets. Notably, in vinculin KO cell sheets, α18-antigen signals were strongly detected at the distorted TJ regions and were significantly increased compared with those in WT epithelial cell sheets. Right images are magnifications of the boxed regions in the left images. Representative images from three independent experiments are shown. Scale bars: 10 μm. (B) The signal intensity of immunostained α18 antibody normalized by the signal intensity of immunostained α-catenin antibody was measured at the bicellular points and tricellular points both in WT and in vinculin KO Eph4 cell sheets. The results of the representative images of three independent experiments are shown. The signal intensity of recorded images was measured without adjustments of brightness and contrast. In vinculin KO cell sheets, the signal intensity of α18 normalized by α-catenin was higher in the tricellular points than in the bicellular points. (C) Confocal super-resolution immunofluorescence images of Myosin-ⅡB, an F-actin marker phalloidin, and a TJ protein ZO-1 in WT (upper) and vinculin KO (lower) Eph4 epithelial cell sheets treated with blebbistatin, an actomyosin tension-relaxing reagent, or DMSO. In the vinculin KO cell sheets treated with 100 μM blebbistatin, the expanded and distorted distribution patterns of myosin IIB (green), phalloidin (red), and ZO-1 (blue) were recovered to linear distribution patterns along cell–cell junctions. Right images are magnifications of the boxed regions in the left images. Representative images from three independent experiments are shown. Scale bars: 10 μm. (D) Confocal super-resolution immunofluorescence images of α18-antigen, which indicates mechanical fluctuation, in WT (left) and vinculin KO (right) Eph4 epithelial cell sheets. Eph4 epithelial cell sheets treated with blebbistatin (lower) or DMSO (upper). The α18 antigen signals were concentrated at tricellular regions, as shown by a TJ protein ZO-1 (red), in WT and vinculin KO epithelial cell sheets in DMSO control. The α18-antigen signals were significantly decreased both in WT and in vinculin KO cells (lower). Representative images from three independent experiments are shown. Scale bars: 10 μm.
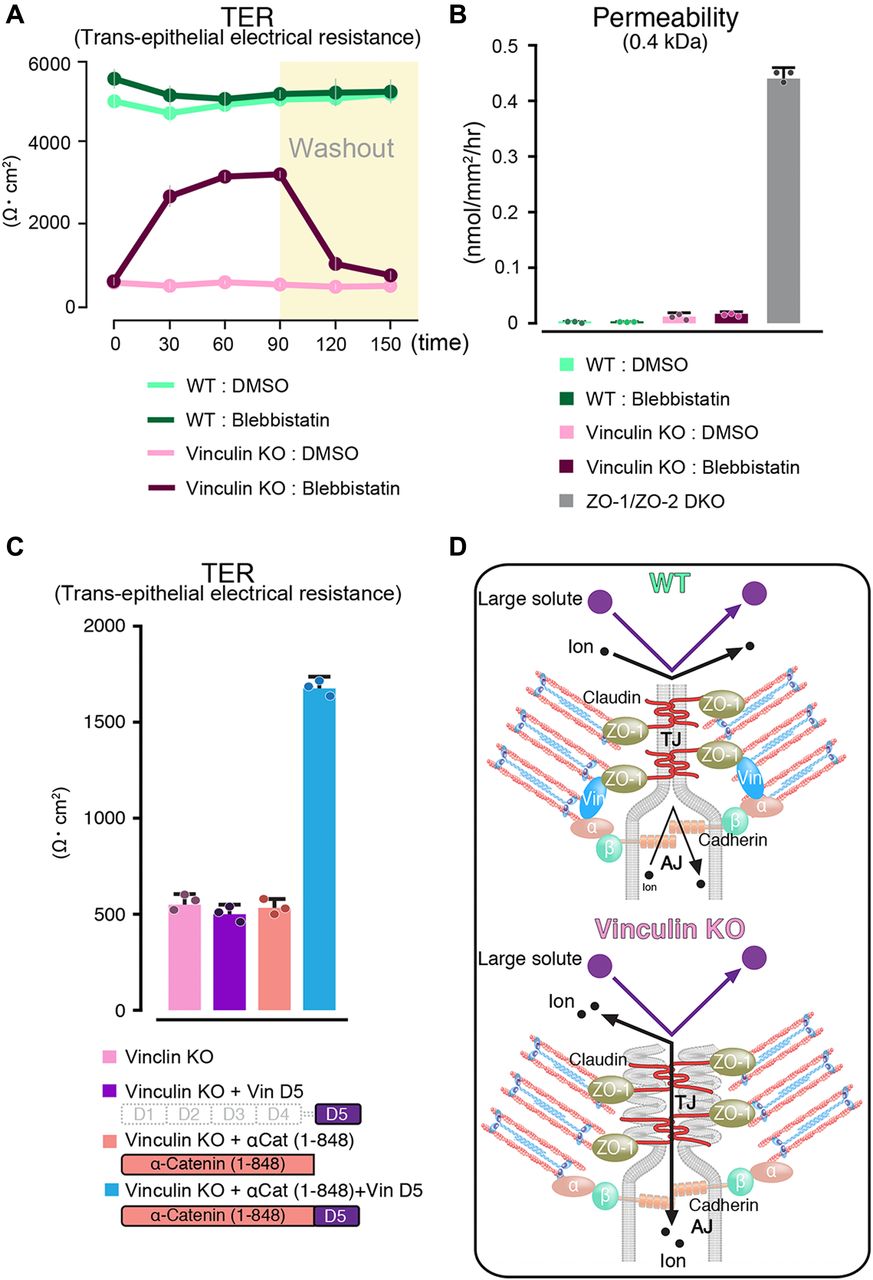
- Figure 5. Mechanical fluctuation induces the loss of the paracellular barrier function for ions in vinculin KO epithelial cell sheets.
(A) The paracellular barrier function for ions, shown by the TER of WT and vinculin KO Eph4 epithelial cell sheets treated with blebbistatin, an actomyosin tension-relaxing reagent, or DMSO. Under the condition of full confluency (13 d of culture on a filter), blebbistatin (100 μM) but not DMSO significantly rescued the TER reduction in the vinculin KO epithelial cell sheets. Results from three distinct clones are shown as the mean ± SEM (n = 3/group). (B) Paracellular barrier function for large solutes, shown by paracellular tracer flux assays using 0.4-kD fluorescein, in WT and vinculin KO Eph4 epithelial cell sheets treated with blebbistatin, an actomyosin tension-relaxing reagent, or DMSO and in ZO-1/ZO-2 DKO Eph4 epithelial cell sheets. Under the condition of full confluency (13 d of culture on a filter), the slight increase in the paracellular permeability for 0.3-kD fluorescein in the vinculin KO epithelial cell sheets was not restored by blebbistatin (100 μM). Results from three distinct clones are shown as the mean ± SEM (n = 3/group). (C) Scheme of the molecules used to transfect vinculin KO Eph4 cells and the TER measured in the monolayers of filter-grown vinculin KO cells and vinculin KO stable clones after transfection. (D) Schematic images showing the AJC as a unified system that maintains the TJ’s paracellular barrier function for ions. Vinculin at the AJ fine-tunes the mechanical fluctuations to maintain the robust, linear distribution of the AJC, which is indispensable for the TJ’s paracellular barrier function for ions. In this mechanism, vinculin at the AJ may be connected with the TJ via ZO-1 to fine-tune the mechanical fluctuation at the AJC. α, α-catenin; β, β-catenin.





Supplementary Materials
- Video 1
Time lapse movie of Venus–occludin-expressing wildtype Eph4 cells.Download video
- Video 2
Time lapse movie of Venus–occludin-expressing vinculin KO Eph4 cells. The movement of occludin–Venus Eph4 cells was registered for 30 min for long-term recording. Venus–occludin-expressing vinculin KO epithelial cells showed much more movement in the cell–cell border than the Venus–occludin-expressing WT epithelial cells. We noted stochastic transitions between the tightly linear and expanded distorted pattern of Venus–occludin in the vinculin KO epithelial cell sheets.Download video




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
