



Article Figures & Data
Figures
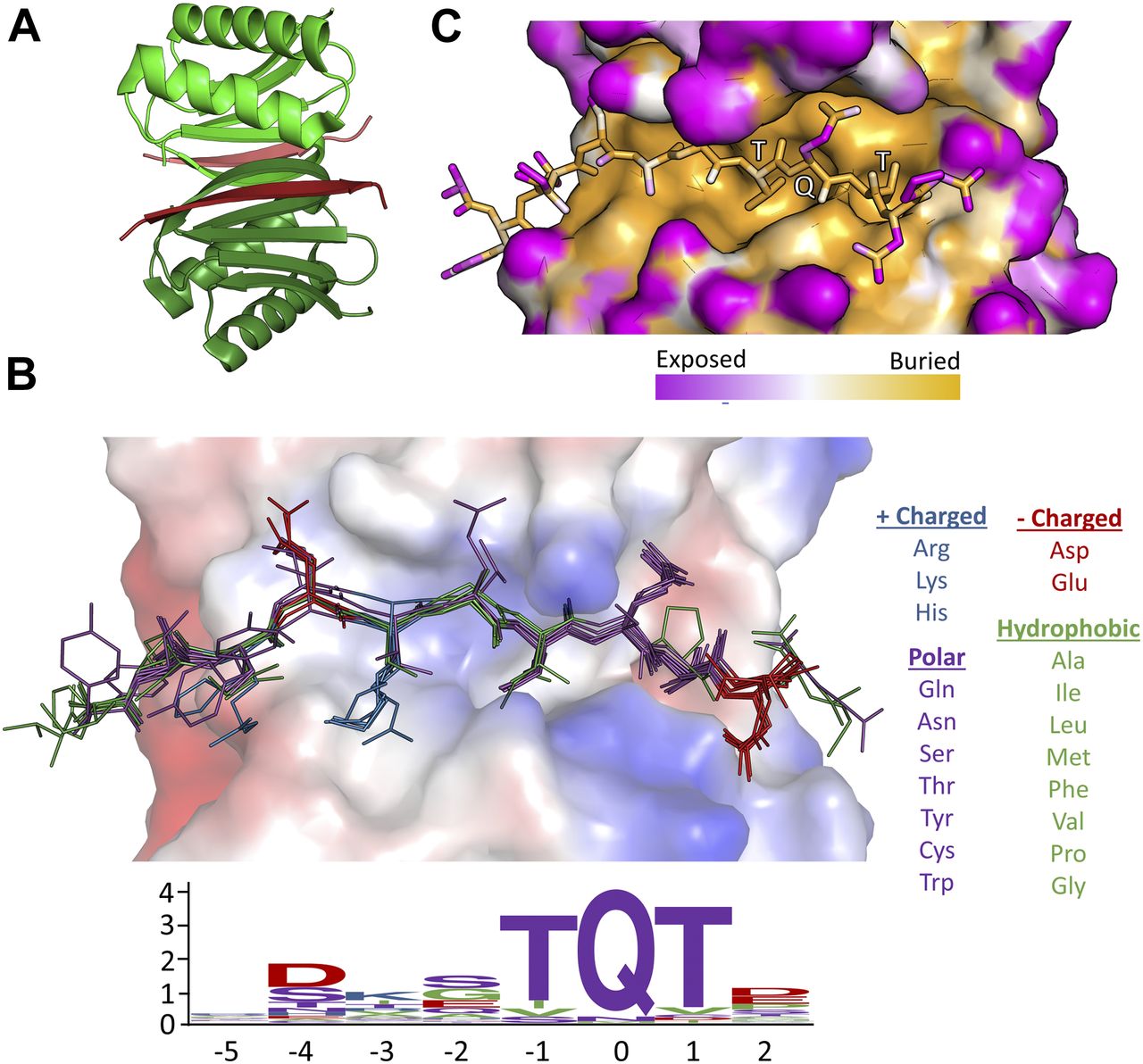
- Figure 1. Motif sequence logo and surface analysis of LC8.
(A) Crystal structure of a representative LC8 dimer (protomers shown in shades of green) bound to a peptide (shades of red). (B) Electrostatic charge potential for the LC8 pocket structure using PyMOL’s charge-smoothed potential calculator, with positive potentials shown in blue, negative in red, and neutral in white. Peptides from available crystal structures of bound LC8 are shown, and colored based upon amino acid chemical characteristics (right). Amino acid enrichment is shown below each position within the LC8-binding motif, calculated from 79 known binder motifs listed on the LC8 database (http://lc8hub.cgrb.oregonstate.edu). Amino acid letter heights represent relative enrichment of that amino acid. (C) Solvent accessible surface area depiction of the same LC8/peptide pair shown in (A). Color scheme was defined at the atomic level using the GetArea program (Fraczkiewicz & Braun, 1998), with magenta representing more solvent exposed and orange regions more buried atoms.
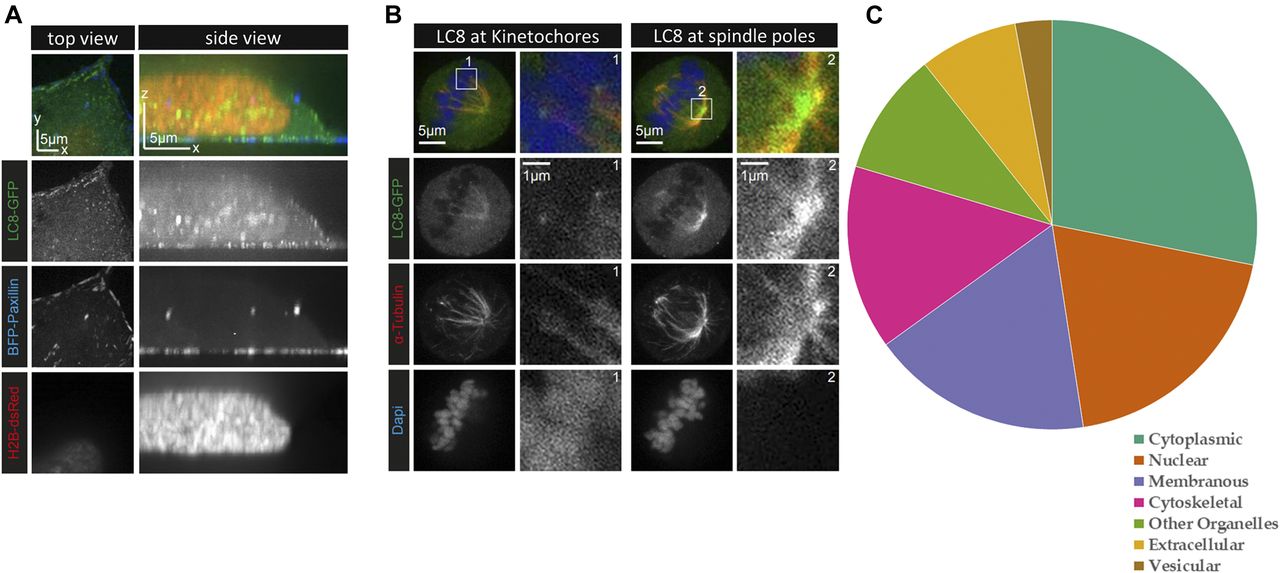
- Figure 2. LC8 and its binding partners display broad cellular localization.
(A) Live HeLa cells stably expressing LC8-GFP (green) were transiently transfected with the focal adhesion marker TagBFP-paxillin (blue) and the nucleus marker dsRed-tagged histone H2B (red). The top view images shown on the left represent an optical section located next to the coverslip. LC8-GFP is present throughout the cell but forms puncta at the cell cortex. (B) HeLa cells stably expressing LC8-GFP were fixed with PFA and stained for the endogenous α-tubulin (red) and with DAPI (blue) to visualize the DNA. LC8 accumulates at the kinetochores (box 1) and at the spindle poles (box 2). In both (A) and (B), images were acquired from a single cell each, using confocal spinning disc microscopy. (C) Localization information derived from the COMPARTMENTS program demonstrates that LC8 binding partners are localized to all cellular compartments. High confidence localization data were available for 59 of the 73 eukaryotic proteins listed on the LC8Hub database.
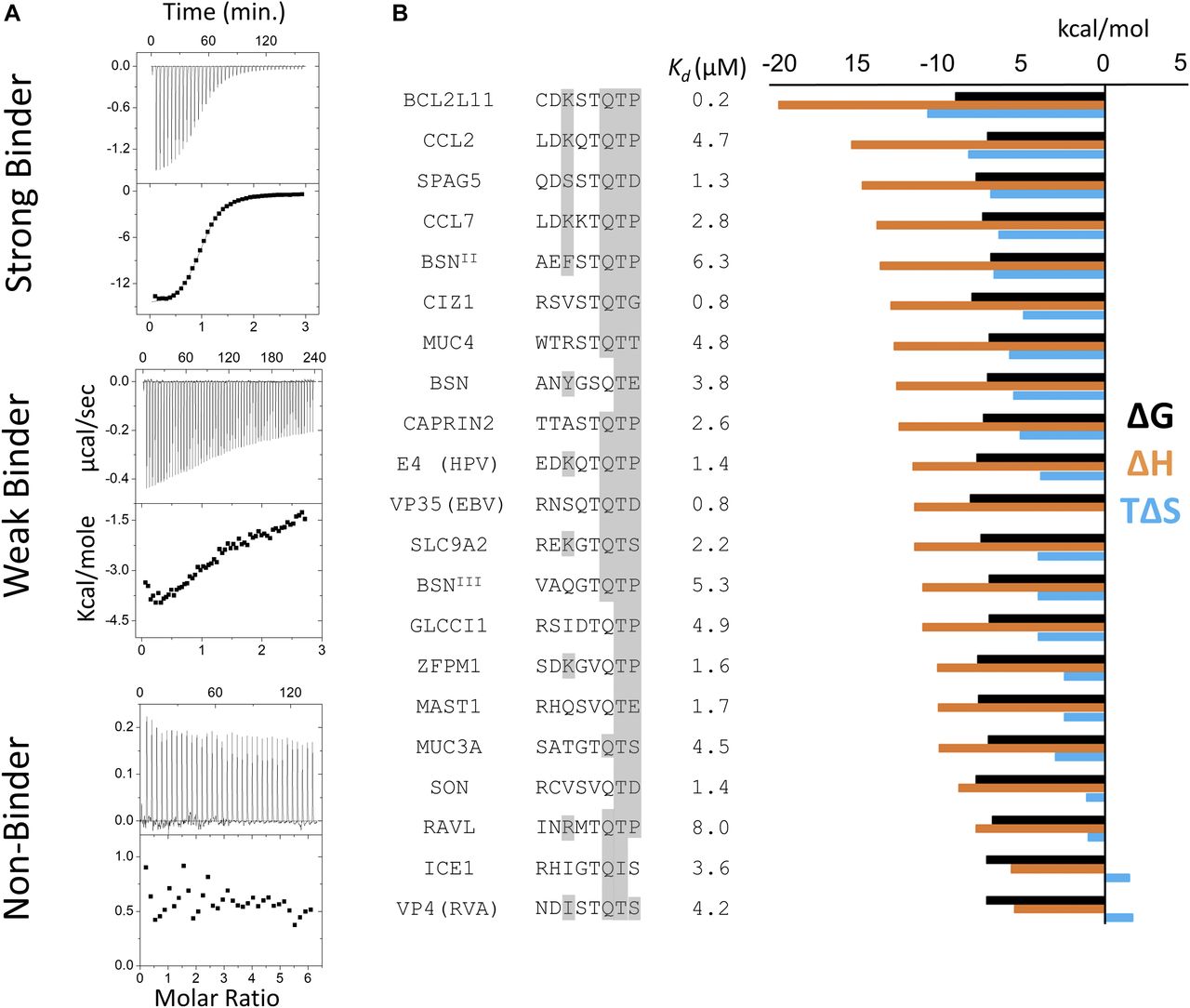
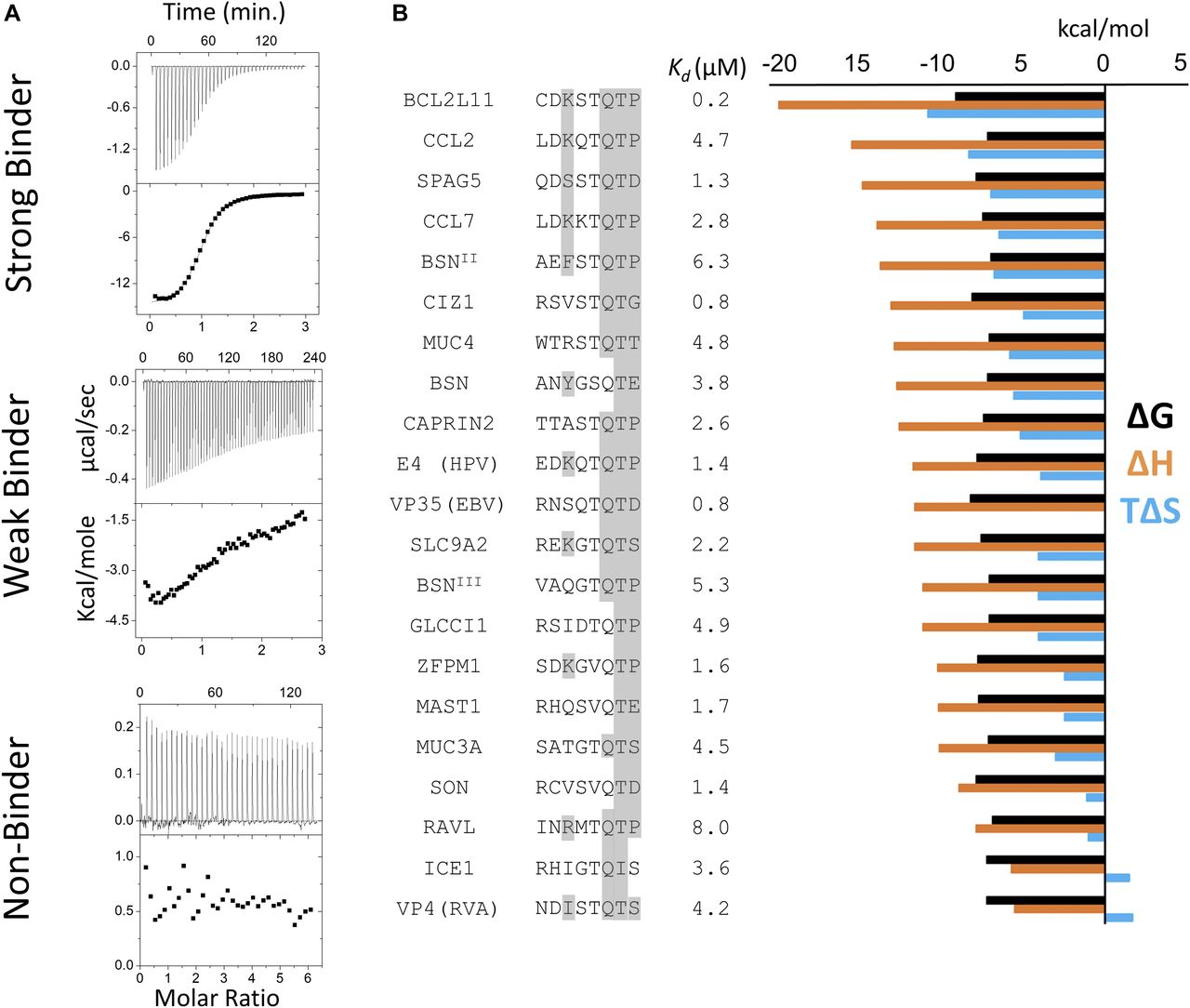
- Figure 3. Thermodynamic analysis of binding peptides.
(A) Representative ITC thermograms of LC8 bound to 14–amino acid–long synthetic peptides. The data were collected at 25°C in 50 mM NaCl, 50 mM NaPO4, and 1 mM NaN3, pH 7.5. Experiments were performed in triplicate. We categorized peptides as strong binders (Kd reliably determined; SPAG5), weak binders (heat generated, but unable to fit the data; TOGARAM1), and nonbinders (QSER1). Weak binders are those with affinities >25 μM. (B) Binding affinities and thermodynamic parameters for strong LC8 binders identified in this study. Thermodynamic parameters for all binding peptides at 25°C are shown. ΔG (black), ΔH (orange), and TΔS (blue) kcal/mol values are the average of two to three independent ITC experiments. Kds are shown in μM. 8 amino acid motifs are shown, with residues capable of making conserved hydrogen bonding interactions highlighted in grey. Sequences are ordered by descending ΔH values.
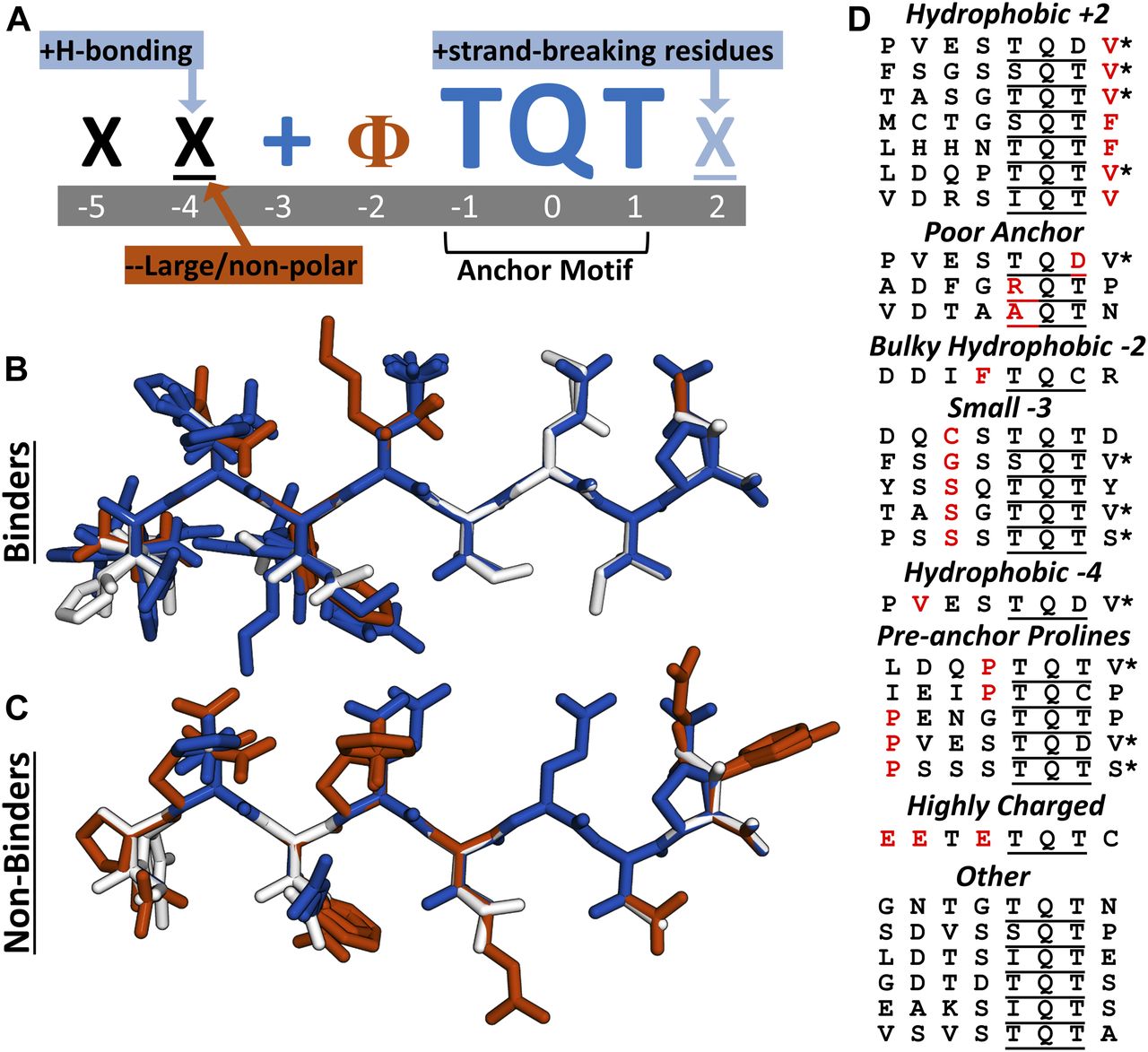
- Figure 4. Analysis of LC8-binding and nonbinding motifs reveals distinct positional preferences.
(A) Motif preferences for LC8 binding partners. “Ф” denotes hydrophobic residues; “X” signifies any residue (unless certain residues are disfavored); underlined “X” signifies any residue but with strong preferences for particular residues; “+” denotes positively charged amino acids. Physiochemical properties beneficial for binding are colored dark blue or light blue, based on magnitude, and deleterious properties are colored in red. (B) All known tightly binding sequences (Kd < 10 μM) are cropped to 8 amino acid motifs and built using the Chimera molecular modeling software. This includes LC8 sequences found on the LC8Hub database, and those determined in this article. (C) Overlay of all nonbinding peptides used in this study. Residues are colored based upon whether they are beneficial (blue), deleterious (red), or neutral (white) for binding, using the amino acid enrichment and depletion in known motifs (Fig 6A). (D) Categories of nonbinding sequences. Residues highlighted in red depict the reason the sequence is placed within a given category. *Denotes sequences placed in multiple categories.
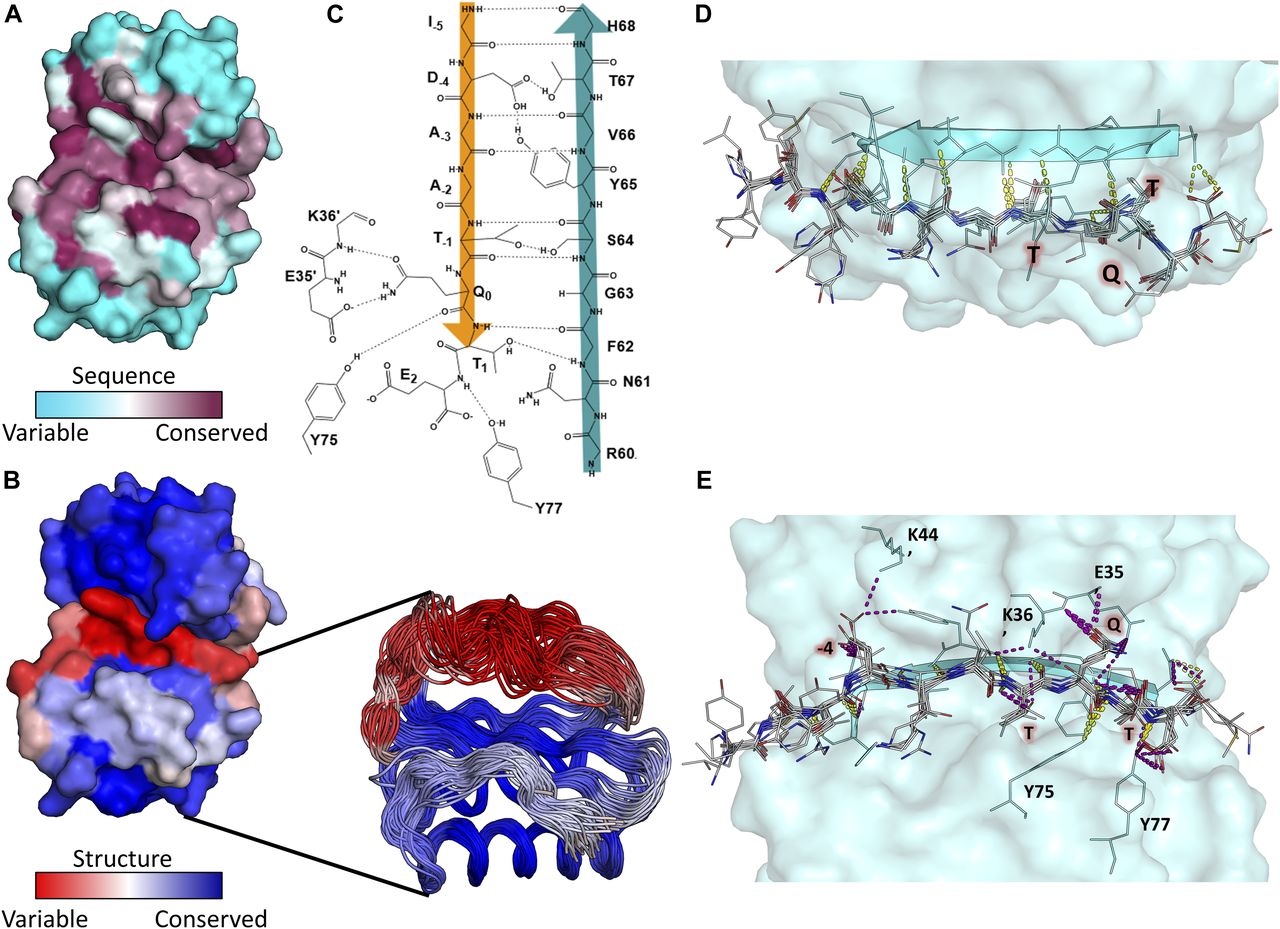
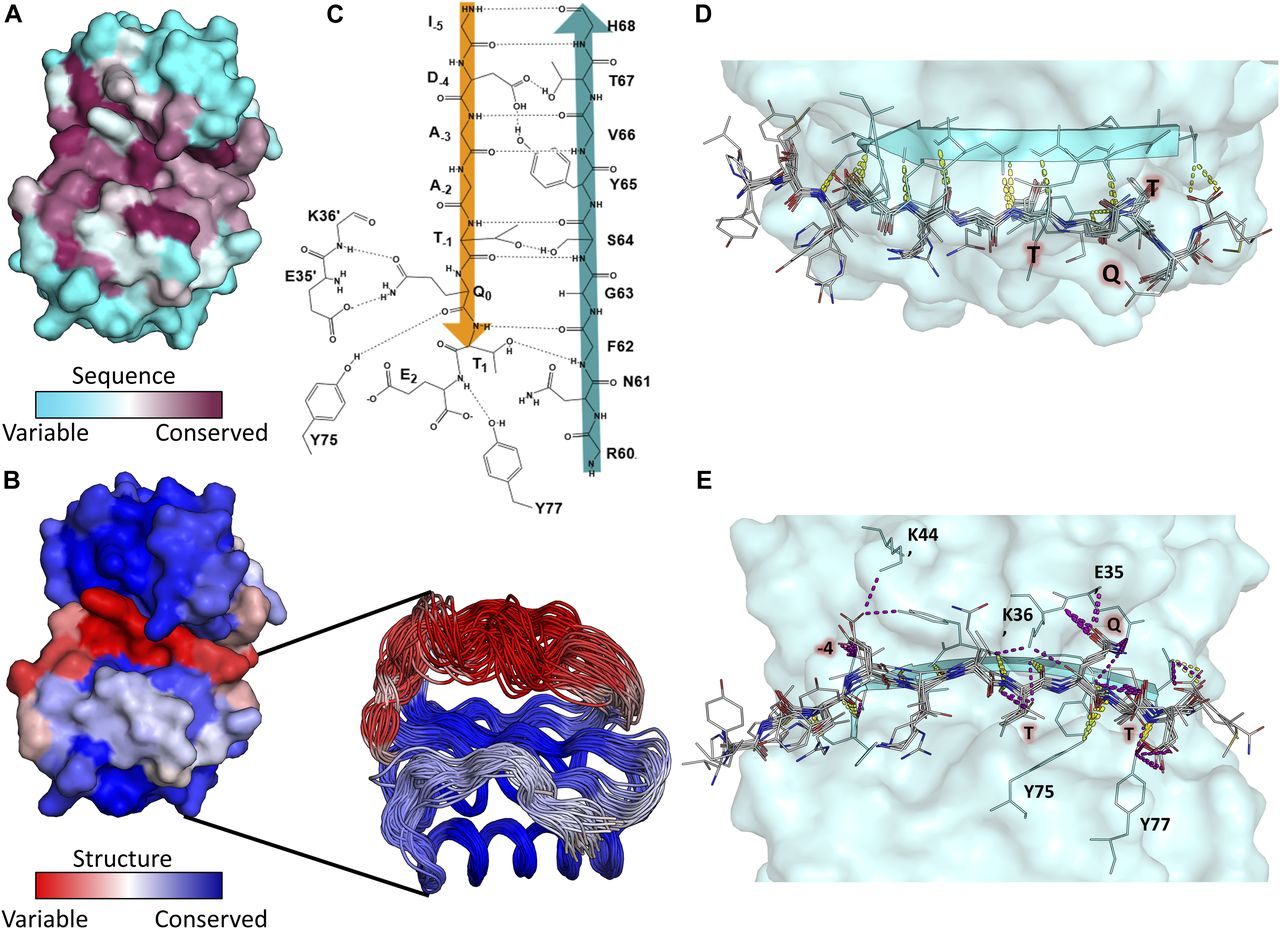
- Figure 5. LC8 is structurally variable but conserved in sequence.
(A) Surface representation of LC8 colored by sequence conservation using ConSurf. More sequence-conserved regions are shown in magenta, less sequence-conserved regions are shown in cyan. Highly conserved residues map to those within the LC8 binding site. (B) Surface representation of LC8 colored by structural conservation in the free protein using the Ensemblator. Regions that are more structurally variable are shown in red, whereas more structurally conserved regions are shown in blue. An overlay of NMR and crystal structure protomers used for the structural analysis is shown as a cut-out in (B). (C) 2D depiction of the binding interface between an example peptide (orange) and the binding β-strand within LC8 (Teal). (D, E) Polar bonds between LC8 and peptides from crystal structures are shown in (D) (top down view, only backbone interactions) and (E) (pocket view). Colors of polar contacts are based on whether the polar contacts stem from backbone (yellow) or side chain (purple) residues on the peptide. Peptide residues with frequent side chain interactions are labeled in red. (C, E) Residues outside of the binding β-strand that are important interaction sites shown in (C) are labeled in (E).
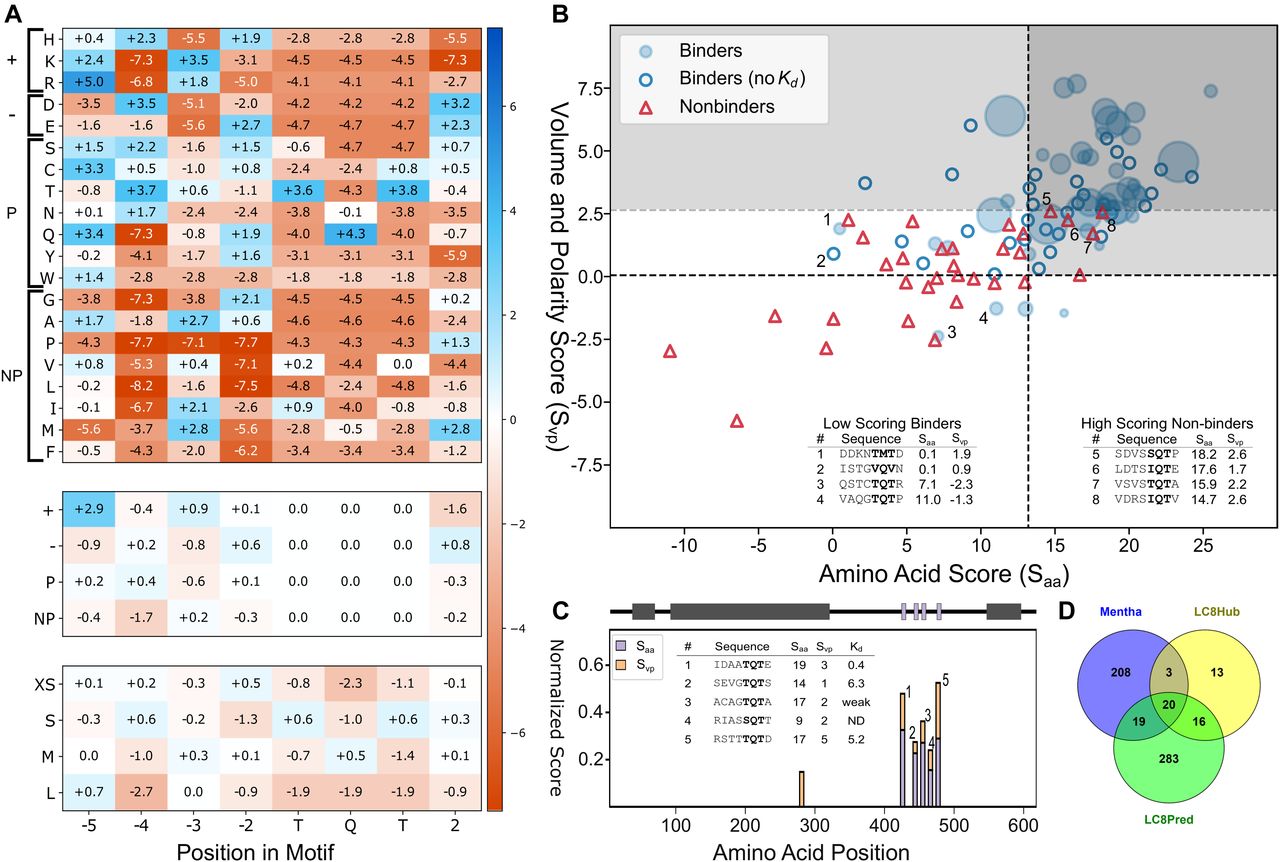
- Figure 6. Generation and testing of The LC8Pred algorithm.
(A) PSSMs for amino acids (A, top), bins by chemical property—positively charged, negatively charged, polar, or nonpolar (middle), and bins by volume—less than 106 A3, 122 to 142 A3, 155 to 171 A3, and greater than 200 A3 (bottom). Values correspond to the combined weight at a given position for the binder-only matrix and the nonbinder-normalized matrix. (B) Scatterplot of available sequences scored using a leave-one-out method of cross validation. For binders with a known Kd, the size of the bubble was varied inversely with the Kd, with binders with a Kd below 0.5 μM represented as the maximum possible dot size. Binder sequences with an unknown binding affinity were plotted as hollow circles and nonbinders as red triangles. The light grey box denotes predicted binding sequences using this scoring system. A second threshold for the volume and polarity axis indicates the very high confidence region, above which the specificity is unity. Outliers are noted in the tables (inset) and numbered in figure. (C) Normalized scores from matrices used to evaluate known LC8-binding protein Chica, where a score of one equates to the ideal amino acids of physicochemical properties at all positions. A sliding window to evaluate Chica for predicted binding sites across the protein was used, with the “0” position within the motif plotted (i.e., at 400, the 0 position is the 400th amino acid within Chica). A diagram of Chica showing secondary structure prediction (grey) and LC8 binding sites (purple) is above, and sequences predicted to bind are on the right, along with their corresponding scores. (D) Venn diagram of human proteins in the LC8Hub database, proteins that contain at least one LC8-binding sequence as determined by LC8Pred, and proteins reported to bind LC8 in the protein–protein interaction database Mentha (Calderone et al, 2013).


Tables
UniProt Gene Sequence Start End Binder? Citation O43521 BCL2L11 APMSCDKSTQTPSP 108 117 Y Puthalakath et al (1999) Q9UPA5 BSNII PRATAEFSTQTPSP 1,498 1,511 Y Fejtova et al (2009) Q86VQ1 GLCCI1 SSSTRSIDTQTPSV 340 353 Y Hutchins et al (2010) Q96R06 SPAG5 HPETQDSSTQTDTS 463 476 Y Schmidt et al (2010) Q6IMN6 CAPRIN2 NQSFTTASTQTPPQ 791 804 Y O75665 OFD1 AKESCNMETQTSST 153 166 Y Boldt et al (2016) Q02505 MUC3A PVLTSATGTQTSPA 1,800 1,813 Y Q9UBY0 SLC9A2 DDHSREKGTQTSGD 749 804 Y Q9Y2F5 ICE1 EKELRHIGTQISSD 181 194 Y Q9ULV3 CIZ1 ARAGRSVSTQTGSM 13 26 Y den Hollander & Kumar (2006) Q99102 MUC4 SQNHWTRSTQTTRE 200 213 Y Q99102 MUC4 DDNHWTRSTQTTRE 200 213 Y P07359 GP1BA GQGAALTTATQTTHLE 566 581 Weaka Q9Y4F4 TOGARAM1 SKTQQTFGSQTECT 788 801 Weaka Q8WWN8 ARAP3 SPSPTGLPTQTPGF 1,514 1,527 Weaka Q01973 ROR1 DDSGGNATTQTTSD 760 774 Weaka Q8NEZ4 KMT2C IVSCVSVSTQTASD 205 218 N Q9UPA5 BSN-shiftb STQTPSPAPASDMP 1,505 1,518 N Q7Z2Z2 EFL1 DERLMCTGSQTFDD 375 386 N Q02817 MUC2 TPTPTPTGTQTPTT 2,000 2,013 N Q9HC84 MUC5B-3 SMATPSSSTQTSGT 2,673 2,686 N Q8TEC5 SH3RF2 TLVSTASGTQTVFP 714 727 N Q9P2G1 ANKIB1 RGDGSDVSSQTPQT 1,065 1,078 N O43526 KCNQ2 DDPMYSSQTQTYGD 370 380 N P14859 POU2F1 ESGDGNTGTQTNGL 13 26 N P35568 IRS1 LPRKVDTAAQTNSR 841 854 N Q2KHR3 QSER1 KTLTFSGSSQTVTP 374 387 N Q99814 EPAS1 TEAKDQCSTQTDFN 509 522 N Q9Y4K1 CRYBG1 RSFVLPVESTQDVSSQ 550 565 N P49862 KLK7 SFRHPGYSTQTHVN 98 111 N Q92904 DAZL TQDDYFKDKRVHHFRRS 272 288 N Q96FV2 SCRN2 VRTLPRFQTQVDRR 342 355 N Q96FV2 SCRN2 DDTLPRFQTQVDRR 344 355 N Q7Z589 EMSY KITFTKPSTQTTNT 261 274 N Q13952 NFYC CLKETLQITQTEVQ 289 302 N Q9HC84 MUC5B-1 TTLPVLTSTATKST 3,049 3,062 N P53350 PLK1 AASLIQKMLQTDPTAR 278 293 N Q92499 DDX1 DDHSGNAQVTQTKFD 271 282 N Q8NBH2 KY ITSYNSQGTQLTVE 81 94 N Q06190 PPP2R3A LQETLTTSSQANLS 625 638 N Q13618 CUL3 KHSGRQLTLQHHMG 542 555 N Q9H4B6 SAV1 NQSFLRTPIQRTPH 70 83 N Q2TV78 MSTL1 EGYRGTANTTTAAYLA 259 274 N Q6ZU65 UBN2 PLQATISKSQTNPV 942 955 N Q96SC8 DMRTA2 SSRSAFSPLQPNAS 433 446 N Q6ZRI0 OTOG TLQQPLELTASQLPAG 1,541 1,556 N Q96JG9 ZNF469 RAAALPEETRSSRR 1,014 1,027 N Anchor motifs are underlined.
Aspartates shown in italics were added to increase solubility.
↵a Peptides that displayed an interaction with LC8 via ITC, but the data were not of sufficient quality to obtain reliable Kd measurements.
↵b BSN-ProP-PD is the Bassoon sequence pulled down by phage display, without shifting the TQT sequence into the correct position.
UniProt Gene Sequence Start End Binder? Citation Q6LCS3 E4 (HPV) YLQGRQEDKQTQTPPP 16 30 Y Q8IX07 ZFPM1 PAPPSYSDKGVQTPSK 947 962 Y P11193 VP4 (Rotavirus A) YVTNSLNDISTQTSTI 600 614 Y P13500 CCL2 YDSMDHLDKQTQTPKT 85 99 Y P18583 SON YSRKSRCVSVQTDPT 87 100 Y Q9Y2H9 MAST1 YGCTRHQSVQTEDG 1,387 1,399 Y Navarro-Lérida et al (2004) P80098 CCL7 QDFMKHLDKKTQTPKL 84 99 Y Q5K4E3 PRSS36 YGPDGEETETQTCPP 468 581 N P20702 ITGAX YGQIAPENGTQTPSP 1,146 1,159 N P03586 MT/HEL (TMV) AQPKQKLDTSIQTEYP 1,305 1,320 N Q8IYH5 ZZZ3 KSVAENGDTDTQTSMF 237 252 N Q5DMI6 DNLJ2 (phage T5) YKIEIPTQCPSCGSK 2 15 N Q92904 DAZL YPQKKSVDRSIQTVVS 243 257 N Q9NZ56 FMN2 YHHRILEAKSIQTSPT 735 749 N Q13418 ILK MDDIFTQCREGN 1 12 N O43432 EIF4G3 DFTPAFADFGRQTPGG 676 691 N Q99613 EIF3C YELMASLDQPTQTVVM 830 844 N O15444 CCL25 NKVFAKLHHNTQTFQA 94 109 N P20042 EIF2S2 KPFMLDEEGDTQTEET 21 36 N Anchor motifs are underlined.
Tyrosines shown in italics were added for accurate concentration determination.
Supplemental Data 1.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- Entering new circles: Expansion of the LC8/DYNLL1 interactome in the ciliary-centrosomal network through system-driven motif evolution
- Quantifying cooperative multisite binding in the hub protein LC8 through Bayesian inference
- Multivalency, autoinhibition, and protein disorder in the regulation of interactions of dynein intermediate chain with dynactin and the nuclear distribution protein
- Native proline-rich motifs exploit sequence context to target actin-remodeling Ena/VASP proteins
- Molecular principles of Piwi-mediated cotranscriptional silencing through the dimeric SFiNX complex
- Dimerisation of the PICTS complex via LC8/Cut-up drives co-transcriptional transposon silencing in Drosophila
- The molecular principles of Piwi-mediated co-transcriptional silencing through the dimeric SFiNX complex
- The dynein light chain 8 (LC8) binds predominantly "in-register" to a multivalent intrinsically disordered partner