



Article Figures & Data
Figures
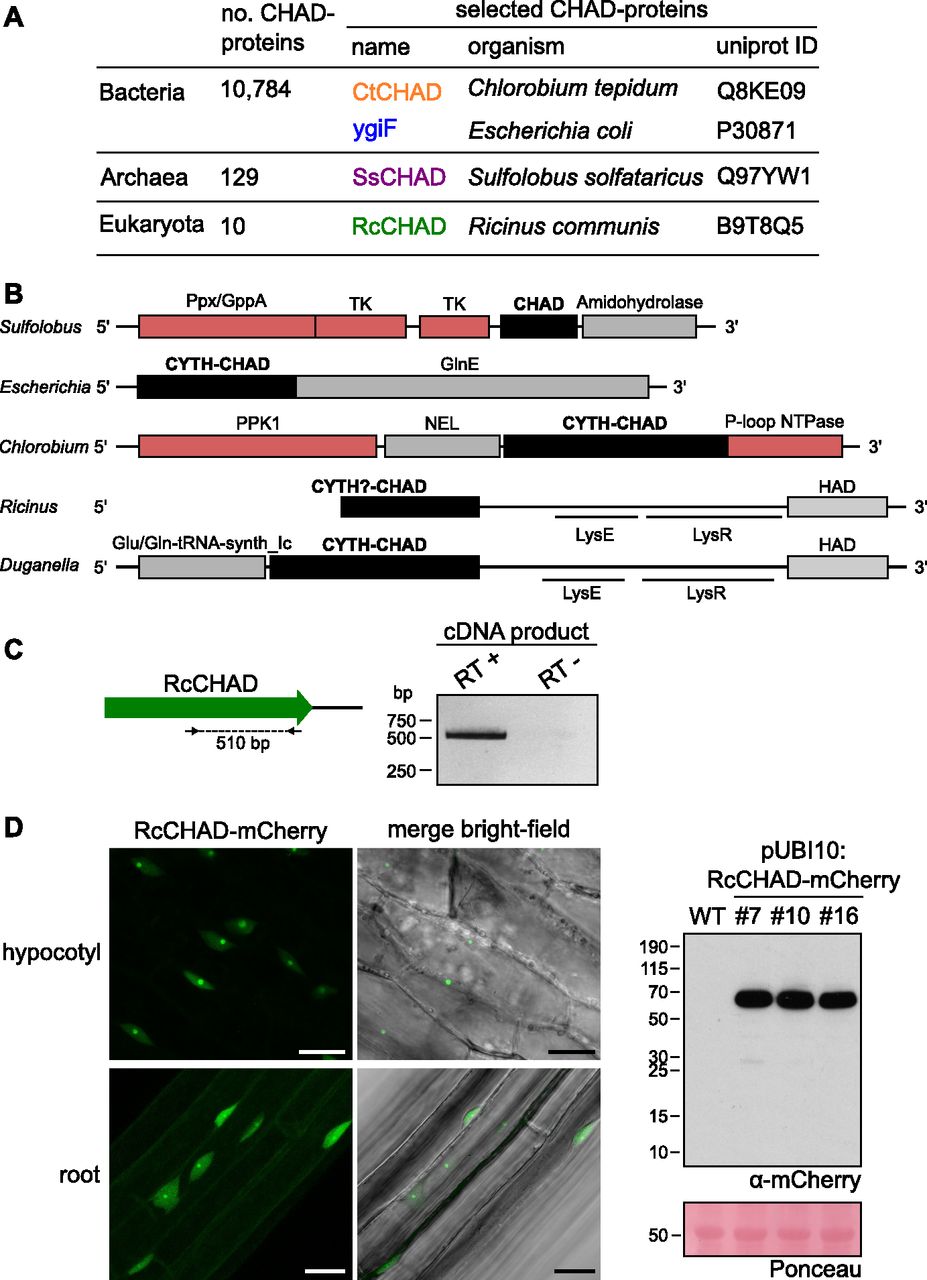
- Figure 1. CHAD domain–containing proteins are present in all kingdoms of life.
(A) Overview of CHAD domain–containing proteins characterized in this study. (B) Genetic loci of genes encoding CHAD domain–containing proteins (in black). Genes coding for polyP-metabolizing enzymes are highlighted in red. The DNA sequence upstream of RcCHAD is missing from contig RCOM_0386220 in the NCBI database. (C) RT-PCR using primers binding to the RcCHAD coding sequence (left) results in a specific product amplified from cDNA from R. communis leaves (right; DNA sequence in Fig S2). (D) Confocal microscopy of transgenic Arabidopsis lines expressing Ubi10p:RcCHAD-mCherry reveals RcCHAD to localize in the nucleus and nucleolus of root and hypocotyl cells (left, scale bars correspond to 20 μm). A Western blot using an anti-mCherry antibody reveals a specific band migrating at the predicted size of the RcCHAD-mCherry fusion protein (63 kD). The Ponceau-stained membrane is shown as loading control below (the major 55-kD band corresponds to RuBisCo).
- Figure S1. Sequence alignment of CHAD domains.
Structure-based sequence alignment of CtCHAD (C. tepidum, UniProt ID Q8KE09), RcCHAD (R. communis, B9T8Q5), DugCHAD (Duganella sp., A0A1H7Y2Q3), ReCHAD (R. eutropha, H16_A0104), ReTTMCHAD (R. eutropha, H16_B1017), and SsCHAD (S. solfataricus, Q97YW1) and including a secondary structure assignment for CtCHAD calculated with DSSP (Kabsch & Sander, 1983). The α-helices belonging to the two 4-helix bundles are colored in dark and light blue, respectively. Histidines involved in Zn2+ ion coordination in the RcCHAD structure are highlighted in yellow, conserved basic residues contributing to the polyP-binding surface are shown in blue, and numbers indicate the positions of residues in CtCHAD used in the mutational analysis presented in Fig 5.
- Figure S2. Sequencing results of the RT-PCR product amplified from Ricinus cDNA using RcCHAD-specific primers.
Primers are shown in green, the obtained sequence matches the NCBI contig ID RCOM_0386220.
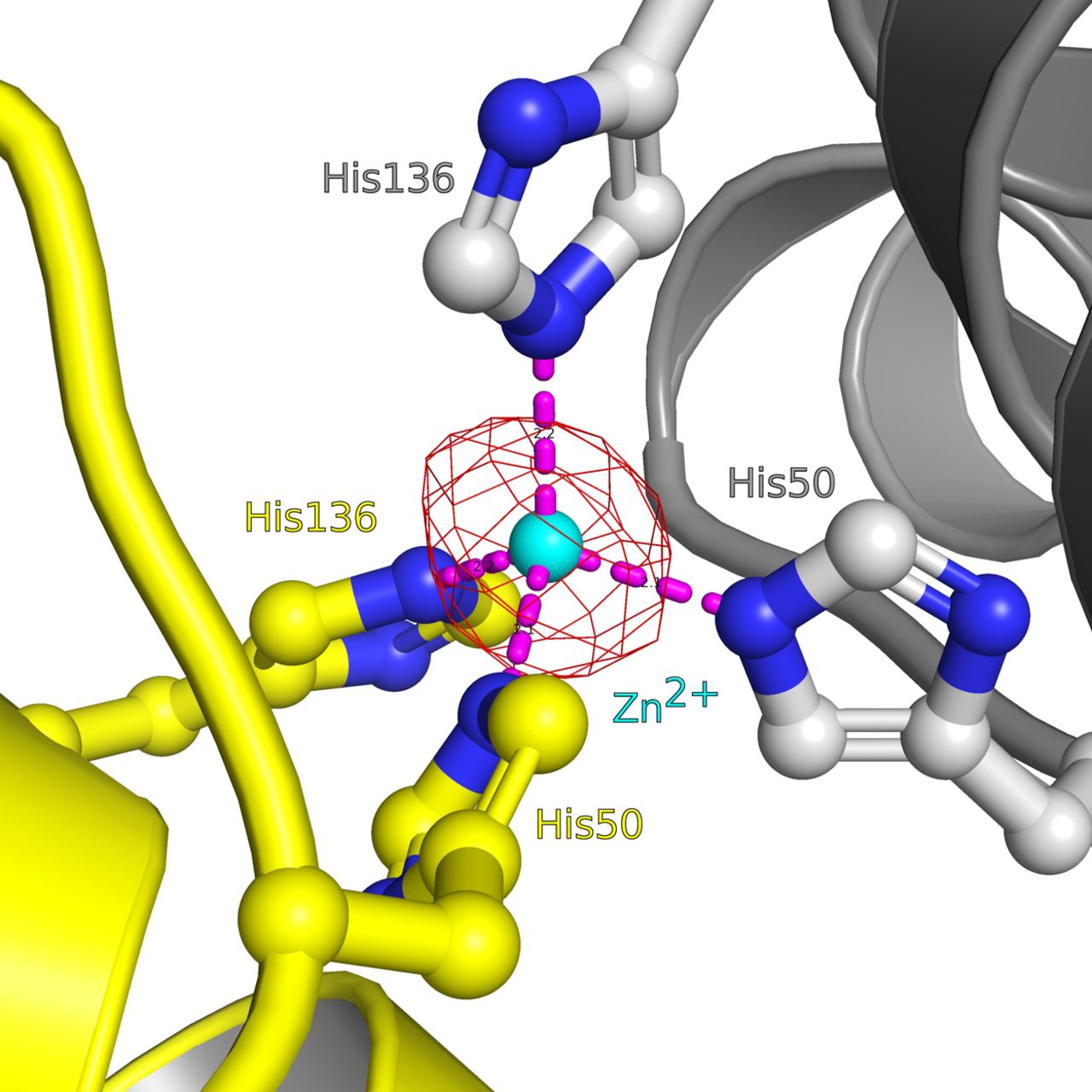
- Figure S3. Close-up view of the Zn2+-binding site located in the RcCHAD structure.
The binding site around the Zn2+ ion (cyan sphere) is formed by the non-conserved His50 and His136 (in bonds representation, interactions shown as dotted lines) at the interface of two symmetry-related molecules (shown in yellow and gray, respectively). A phased omit difference density map contoured at 8 σ is shown alongside (red mesh).
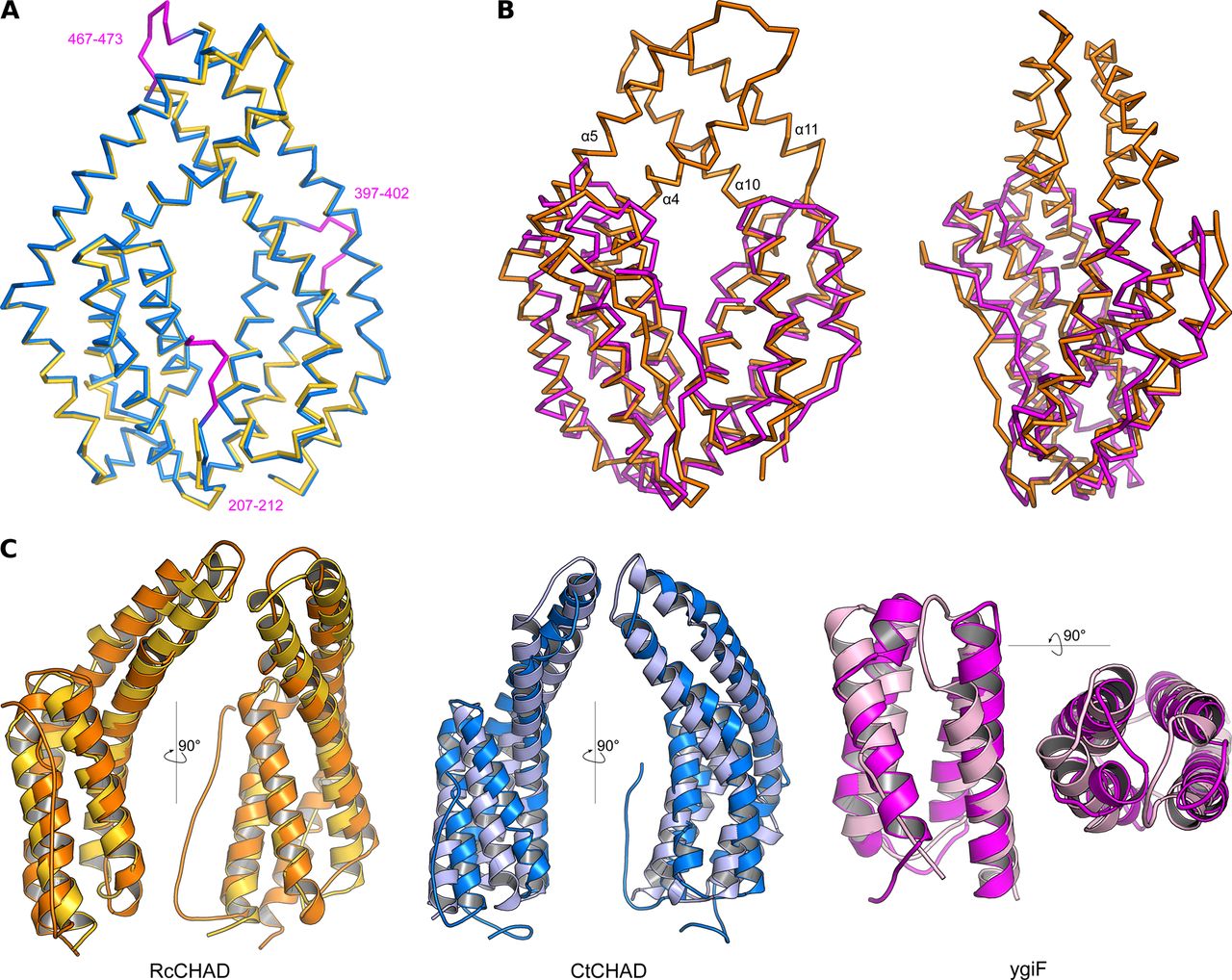
- Figure S4. CHAD domains are formed by two 4-helix bundles related by an internal pseudo twofold symmetry.
(A) Structural superposition of the CtCHAD structure (in blue) with a second CtCHAD crystal form (PDB-ID 3E0S, in yellow; r.m.s.d. is ∼0.6 Å comparing 290 corresponding Cα atoms). Shown are Cα traces, with loop regions present in our structure and missing in PDB-ID 3E0S highlighted in magenta. (B) Structural superposition of RcCHAD (in orange) with the E. coli ygiF CHAD domain (PDB-ID 5A61, residues 205–428, in magenta) reveals the presence of protruding helices in RcCHAD forming the central pore (r.m.s.d. is ∼2.4 Å comparing 184 corresponding Cα atoms). (C) Structural superposition of the two 4-helix bundles in RcCHAD (residues: 7–166 versus 167–303), CtCHAD (residues: 207–373 versus 374–522), and ygiF (residues: 205–336 versus 337–428) shown as ribbon diagrams in two orientations.
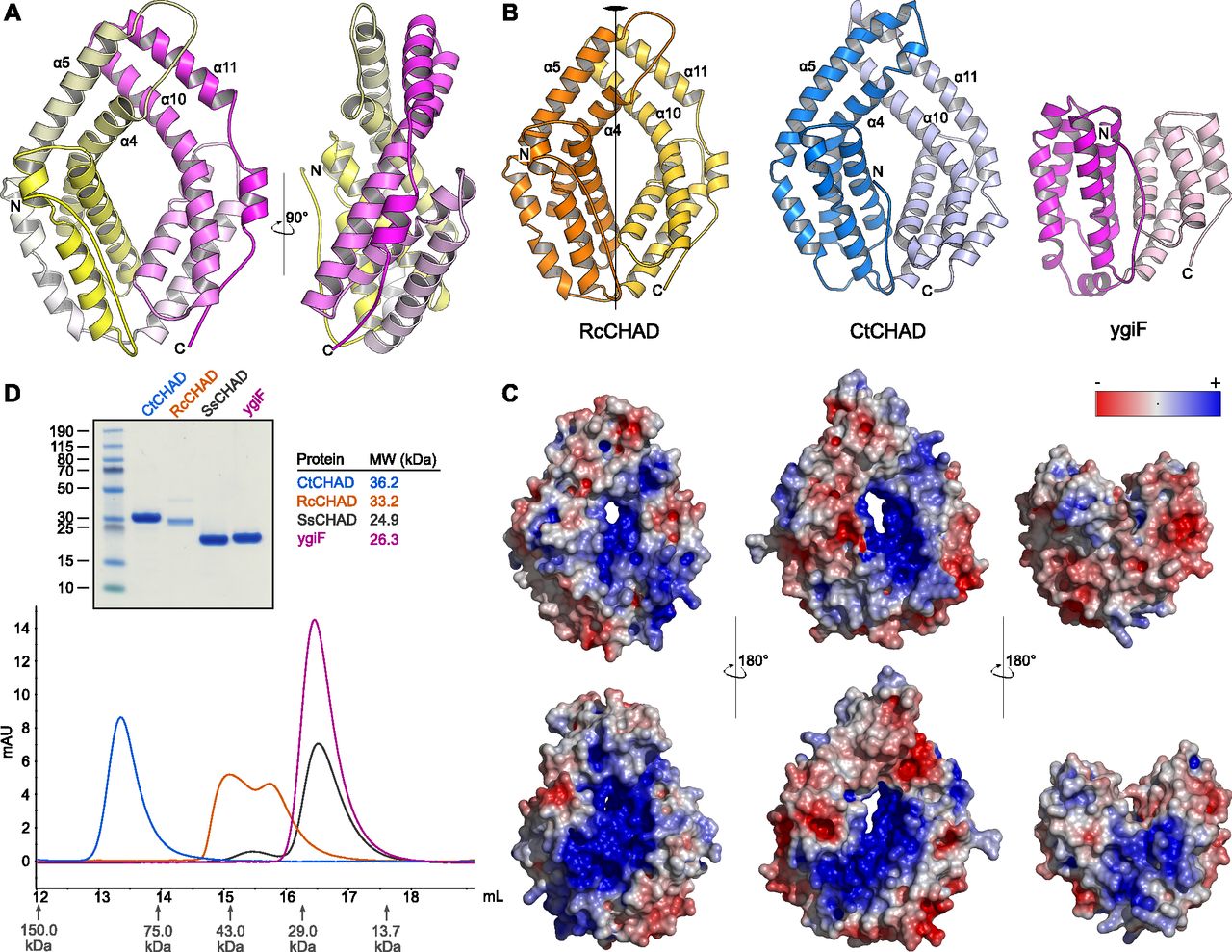
- Figure 2. CHAD domains are helical bundles with twofold internal symmetry and a conserved basic surface area.
(A) Architecture of the CHAD domain. Shown is a ribbon diagram of RcCHAD colored from N- (yellow) to C-terminus (magenta). (B) Structural comparison of the RcCHAD, CtCHAD, and ygiF CHAD domain (Martinez et al, 2015) structures reveal the presence of two 4-helix bundles in all CHAD domains, related by pseudo twofold symmetry (indicated by a vertical line). Note that the helices α4, α6, α10, and α11 in ygiF are much shorter when compared with RcCHAD and CtCHAD, and hence ygiF lacks the central pore. (C) Identification of a conserved basic surface area in pro- and eukaryotic CHAD domains. Electrostatic potentials calculated in APBS (Jurrus et al, 2018) were mapped onto CHAD domain molecular surfaces in Pymol. Shown are front (upper panel) and back (lower panel) views. A highly basic surface area covers the front- and back side of the CHAD domain and includes the central pore present in RcCHAD and CtCHAD. (D) Analytical size-exclusion chromatography reveals different oligomeric states for the CHAD domains analyzed in this study. An SDS–PAGE analysis of the respective peak fractions (pooled) is shown alongside.
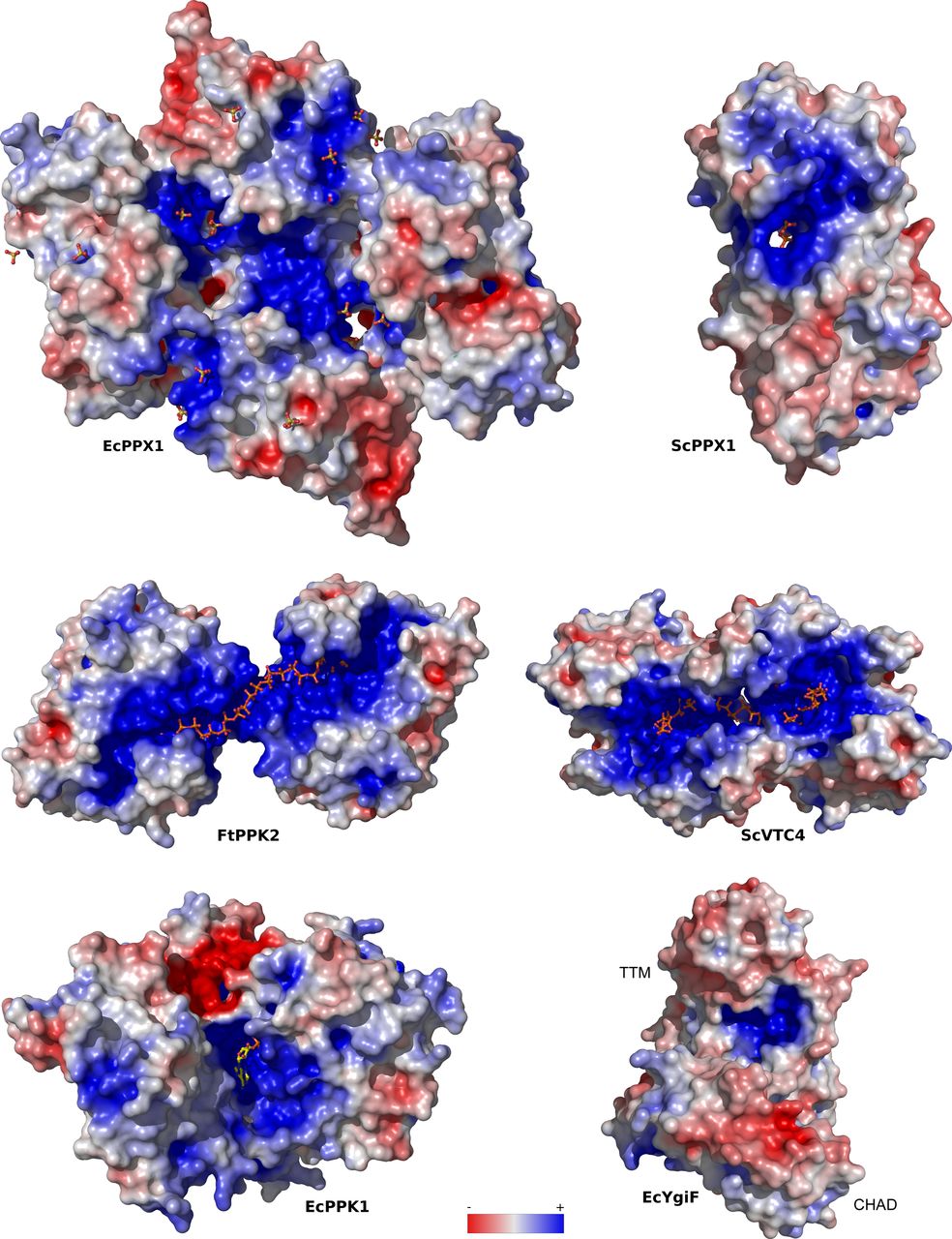
- Figure S5. PolyP-metabolizing enzymes contain large basic surface patches facilitating polyP binding.
Depicted are electrostatic potentials calculated in APBS (Jurrus et al, 2018), mapped onto molecular surfaces calculated in Pymol for EcPPX1 (PDB-ID: 1U6Z), ScPPX1 (PDB-ID: 2QB7), FtPPK2 (PDB-ID: 5LLF), ScVTC4 (PDB-ID: 3G3Q), EcPPK1 (PDB-ID: 1XDP), and ygiF (PDB-ID: 5A61). Bound phosphates, sulfates, polyPs, and PPK1 substrate analogs are highlighted in bond representation.
- Figure S6. Schematic overview of the GCI chip used to determine the interaction between CHAD domain–containing proteins and polyP.
Streptavidin was covalently linked to a 4PCP-WAVE chip (Creoptix AG) and subsequently passivated with BSA (left). Biotinylated polyP (average chain length ∼100 Pi units, right) was then bound to the immobilized strepdavidin on the GCI chip.
- Figure 3. CHAD domains are specific polyP-binding modules.
(A) Quantitative GCI polyP-binding assay. Biotinylated polyP (average chain length is ∼100 Pi units) was immobilized on a streptavidin GCI chip, and the different CHAD domains were used as analytes (Fig S6). The yeast polyP polymerase Vtc4 and BSA were used as positive and negative controls, respectively. Shown are recorded sensograms (in red) with the respective fits (in black) and including table summaries of the derived association rate constant (ka), dissociation rate constant (kd), and dissociation constant (KD). (B) Short-chain polyPs (average chain length ∼7 Pi units) can compete with CHAD domains for binding to the polyP-coated GCI chip. Shown are dose–response curves with derived IC50 estimates. (C) Diadenosine pentaphosphate (AP5A) cannot compete with RcCHAD for binding to the polyP-coated GCI chip (right panel) as efficiently as polyP (left panel, average chain length ∼7 Pi units). Shown are sensograms of the association phase at indicated inhibitor concentration. (D) Sensograms for SsCHAD and RcCHAD reveal no significant interaction with biotinylated single-stranded DNA (54 nt, in orange) or single-stranded RNA (10 nt, in green). Biotinylated polyP (average chain length ∼100 Pi units, in red) is shown as a positive control. CtCHAD, however, binds biotinylated heparin. Shown are the recorded sensograms (in red) with the respective fits (in black) and including a table summary of the derived association rate constant (ka), dissociation rate constant (kd), and dissociation constant (KD). (E) Phosphohydrolase activities of ygiF full-length 1–433 (FL) and ygif-TTM1–200 (TTM) versus different phosphorylated substrates. Symbols represent raw data, lines indicate mean values, and error bars denote SD of three independent replicates. An SDS–PAGE analysis of the purified proteins is shown alongside. The theoretical molecular weight is ∼22.3 kD for TTM and ∼48.4 kD for FL.
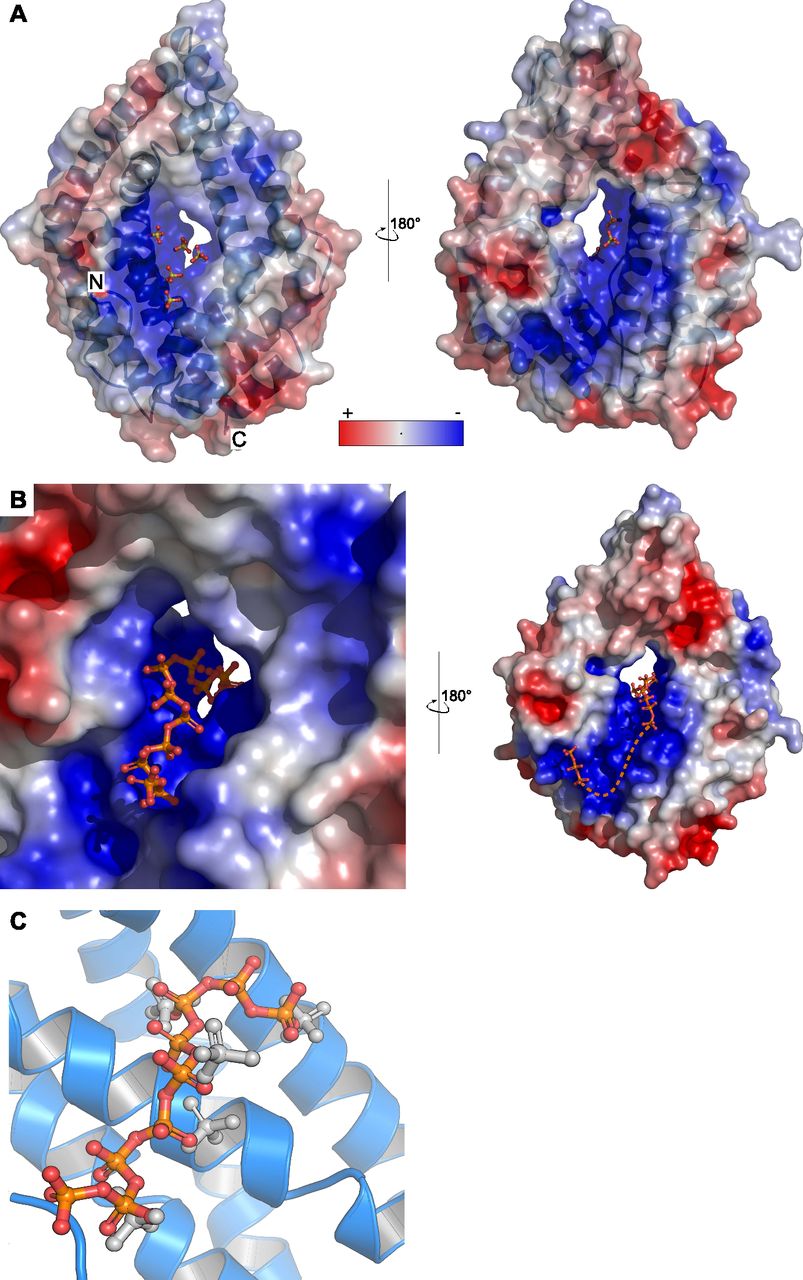
- Figure 4. The basic surface area in CHAD domains provides a binding platform for polyPs.
(A) Sulfate ions (in bonds representation) originating from the crystallization buffer are bound to the basic surface area in CtCHAD. Shown are front and back views of CtCHAD as combined ribbon diagram and molecular surface. An electrostatic potential calculated in APBS was mapped onto the molecular surfaces in Pymol. (B) Overview of the polyP complex structure, obtained by crystallization of CtCHAD in the presence of 5 mM polyP (average length ∼7 Pi units). A polyP 9-mer and a tripolyphosphate moiety could be modeled (in bonds representation), with the polyP 9-mer occupying the central pore and extending to both sides. The dashed line indicates the approximate position of several peaks in the Fo-Fc difference electron density map, which could not be modeled with confidence. (C) Structural superposition of the sulfate ion– and polyP-bound CtCHAD structures (r.m.s.d. is ∼0.5 Å comparing 304 corresponding Cα atoms) reveals that the sulfate ions (in bonds representation, in gray) mimic the position of Pi units in the polyP 9-mer (in orange-red) in the CtCHAD-polyP complex.
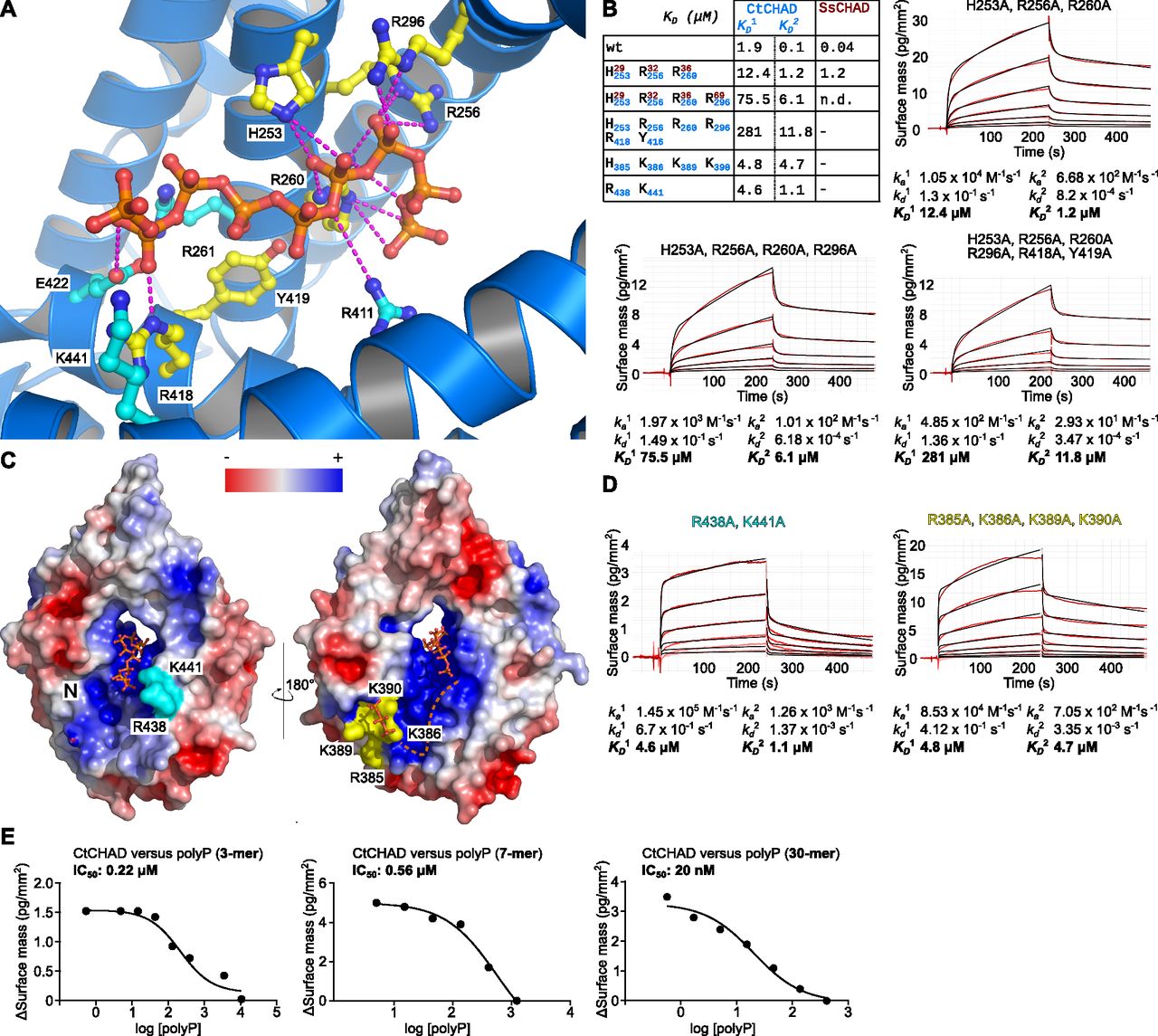
- Figure 5. CtCHAD binds polyP through basic amino-acid residues distributed along the central cavity and the back of the protein.
(A) Detailed view of CtCHAD (blue ribbon diagram) bound to a polyP 9-mer (in orange, in bonds representation) and including selected conserved basic amino acids involved in polyP binding (in cyan, residues included in mutational analyses shown in yellow). (B) Mutations in the central basic binding surface in CtCHAD (dissociation constants in blue; corresponding mutations in SsCHAD in red) strongly decrease polyP binding in GCI assays. Shown are sensograms (in red), the respective fits (in black), and table summaries of the derived association rate constant (ka), dissociation rate constant (kd), and dissociation constant (KD). (C) Identification of two distal polyP-binding surfaces on the front side (shown in cyan) and back side (in yellow) of the CtCHAD—polyP complex structure. (D) Mutations of conserved residues in the two distal surfaces reduce polyP binding as shown in GCI assays. Shown are sensograms (in red), the respective fits (in black), and table summaries of the derived kinetic parameters. (E) GCI competition assays using tripolyphosphate (3 Pi), short-chain (average ∼ 7 Pi units), and long-chain (average ∼ 30 Pi units) polyPs. Shown are dose–response curves with derived IC50 estimates.
- Figure S7. Mutations of conserved basic amino acids in SsCHAD disrupt polyP binding in quantitative GCI assays.
Shown are sensograms (in red), the respective fits (in black), and table summaries of the derived association rate constant (ka), dissociation rate constant (kd), and dissociation constant (KD). Simultaneous mutation of His29 (which corresponds to His253 in CtCHAD), Arg32 (Arg256 in CtCHAD) and Arg36 (Arg260 in CtCHAD) to alanine strongly reduces binding of the mutant protein to polyP (wild-type KD is 44.5 nM). Additional mutation of Arg69 to Ala (Arg296 in CtCHAD) disrupts binding in our GCI assay.
- Figure 6. RcCHAD localizes to the nucleus and nucleolus of tobacco cells and co-localizes with EcPPK1 to EcPPK1-generated polyP granules.
(A) Transient expression of Ubi10p:RcCHAD-mCherry in tobacco leaves reveals a nuclear/nucleolar localization of the fusion protein (top row). Expression of Ubi10p:EcPPK1-mCitrine induces the formation of polyP granules (center row), not observed when using a catalytically impaired version of the enzyme (EcPPK1H435A, H592A, bottom row). Scale bars correspond to 50 μm. Shown are Z-stacks from representative cells from three leaves obtained from three different plants. (B) RcCHAD-mCherry co-localizes with DAPI-stained nuclei and shows a higher intensity in nucleoli (not stained by DAPI). Scale bars correspond to 20 μm. (C) Magnified views of the nuclear localization of RcCHAD-mCherry when expressed in isolation (top row) and its redistribution to EcPPK1-generated polyP granules (bottom row). Scale bars correspond to 20 μm. (D) Western blots using anti-mCherry and anti-mCitrine antibodies reveal that RcCHAD-mCherry (63 kD) and EcPPK1-mCitrine (109 kD) migrated at the expected size in tobacco infiltrated leaves. RuBisCO (detected with Ponceau) is shown below as a loading control.
- Figure S8. PolyP binding may induce oligomerization of CtCHAD.
SEC and RALS analysis of CtCHAD in complex with polyP. (A) RALS traces of CtCHAD in the presence (red) and absence (blue) of 1 mM polyP (average chain length ∼30 Pi units). Indicated oligomeric states are calculated based on CtCHAD, as shown in (B). (B) RALS traces (blue) and extrapolated molecular weight (black) of the two peaks of CtCHAD. Retention volumes and calculated molecular weights are shown alongside. In the absence of polyP, CtCHAD forms a dimer (∼90%) and a tetramer (∼10%). The theoretical molecular weight of monomeric CtCHAD is 36,194 D. (C) Putative tetrameric and dimeric assemblies for CtCHAD based on crystal packing analysis as implemented in the program PISA (Krissinel & Henrick, 2007). Shown are molecular surfaces of a CtCHAD tetramer (left) and dimer (right) in two orientations, respectively. Note that the position of the central cavities in the tetramer and the polyP-binding surfaces in the dimer could allow for the cooperative binding of long-chain polyPs.








Supplementary Materials




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- Ionic polyphosphorylation of histidine repeat proteins by inorganic polyphosphate
- Structural and functional insight into the plant unique multimodular triphosphosphate tunnel metalloenzymes of Arabidopsis thaliana
- In Vitro Analytical Approaches to Study Plant Ligand-Receptor Interactions
- Acidocalcisomes and Polyphosphate Granules Are Different Subcellular Structures in Agrobacterium tumefaciens