



Article Figures & Data
Figures
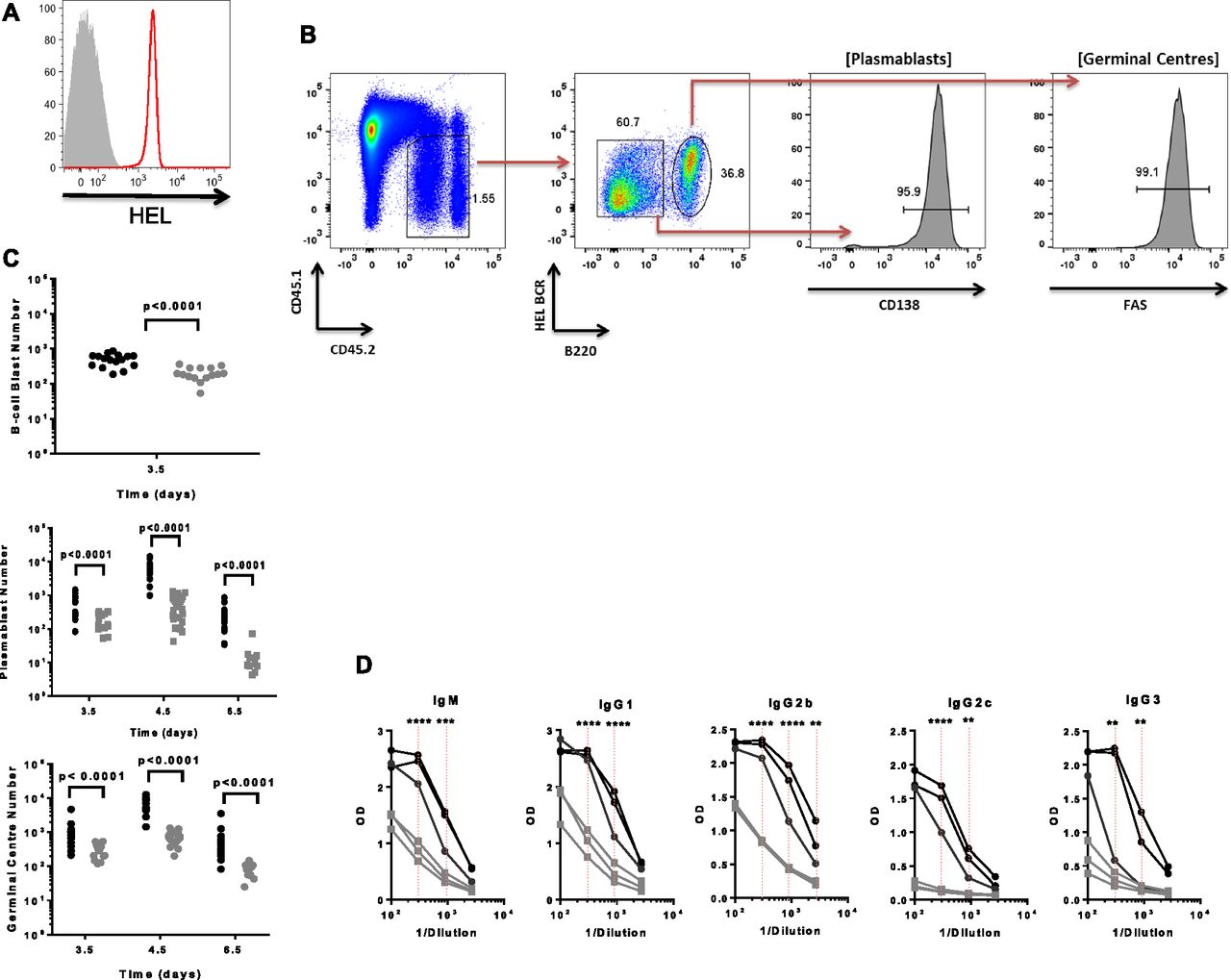
- Figure 1. miR-155 is required to sustain the plasmablast B-cell response.
(A) A representative histogram showing HEL expression level on conjugated HEL-SRBCs (red) compared with unstained control (grey). (B) Representative flow cytometric plot showing gating strategy for SWHEL Mir155+/+ B cells at days 4.5 post immunisation, for identification of CD45.2+ donor derived HEL BCR+, B220lo plasmablast B cells or HEL BCR+, B220hi germinal centre B cells. (C) The number of SWHEL Mir155+/+ (black) or Mir155−/− (grey) HEL-specific B-cell blasts, plasmablast B cells and germinal centre B cells was calculated per 106 lymphocytes after immunisation in mice (N = 16–19 independent Mir155+/+ samples and 10–24 independent Mir155−/− samples). Data are representative of at least two independent experiments. For B-cell blast data, a Welch’s t test was used. For plasmablast and germinal centre data, P-values are from t tests using the error mean square from the ANOVA. (D) HEL-specific antibodies of the indicated immunoglobulins were measured in the serum of mice injected with SWHEL Mir155+/+ (black) or Mir155−/− (grey) B cells, at day 4.5 post immunisation with HEL-SRBCs. Red dotted line represents statistical analysis of indicated Mir155+/+ or Mir155−/− values using two-way ANOVA with Sidak’s multiple comparison test where **P < 0.01, ***P < 0.001, ****P < 0.0001.
- Figure 2. miR-155 is essential for the optimal proliferation of plasmablast B cells.
(A) SWHEL Mir155+/+ (black line) or Mir155−/− (grey line) HEL-binding, B220+ B cell blasts were assessed for different CFSE dilutions. (B) The percentage of CFSE+ cells in each division at days 2.5 and 3.5 post immunisation. The generation number is indicated above each data set. A two-way ANOVA with Sidak’s multiple comparison test was used. Values shown are mean ± SD. (C) Cell cycle analysis using DAPI staining was carried out on SWHEL Mir155+/+ or Mir155−/− plasmablast B cells at 3.5 and 4.5 d post immunisation. The frequency of cells in the S-G2-M phases in Mir155+/+ (black) or Mir155−/− (grey) plasmablasts is shown. A nonparametric Mann–Whitney test was used per time point. (D) Representative flow cytometry plot of DAPI staining at 4.5 d post immunisation in miR-155–sufficient or miR-155–deficient plasmablast B cells. Cells previously gated on lymphocytes and single cells. (E) EdU incorporation in SWHEL Mir155+/+ (black) or Mir155−/− (grey) plasmablast B cells at the time points indicated post immunisation. Two-way ANOVA was used. There was no significant interaction between genotype and day, but both genotype and day had significant effects P < 0.001. (F) Representative flow cytometry histograms of EdU incorporation in wild-type (WT, black line) or miR-155–deficient plasmablast B cells (black line) compared with plasmablast B cells from control mice not injected with EdU (grey) at 4 d post immunisation. Data are representative of at least two independent experiments.
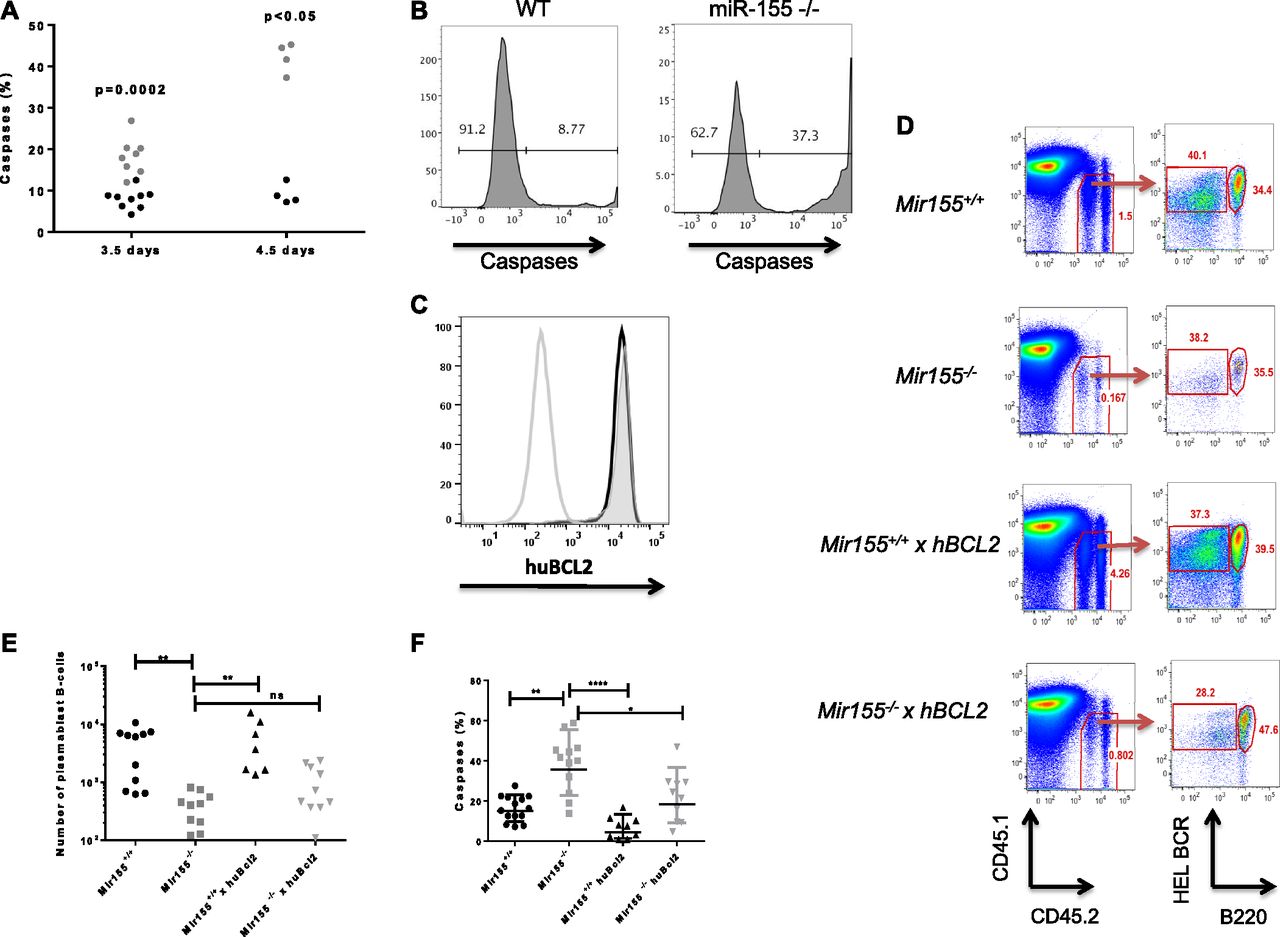
- Figure 3. miR-155 regulates survival of plasmablast B cells.
(A) SWHEL HEL BCR+ B220lo Mir155+/+ (black) or Mir155−/− (grey) plasmablast B cells were analysed for activated caspases at the time points shown. A nonparametric Mann–Whitney test was used per time point. (B) Representative flow cytometry histograms of cleaved caspase detection in wild-type or miR-155−/− SWHEL plasmablast B cells at day 4.5 post immunisation. (C) Expression of the human Bcl2 transgene (huBCL2) in SWHEL Mir155+/+ × Bcl2 (black line) or SWHEL Mir155−/− × Bcl2 (solid grey line) plasmablast B cells, compared with negative control (single grey line). (D) Representative FACS plot showing the gating strategy for analysis of adoptively transferred SWHEL Mir155+/+ or Mir155−/− B cells expressing a human Bcl2 transgene at day 4.5 post immunisation. Plots were previously gated on lymphocytes and single cells and analysed for CD45.2 donor cells and HEL+ B220lo plasmablast B cells. HEL+ B220hi germinal centre B cells could also be visualised in all mouse strains. (E) The number of splenic SWHEL Mir155+/+ or Mir155−/− plasmablast B cells in mice with or without the expression of a human Bcl2 transgene per 106 lymphocytes. A nonparametric Kruskal–Wallis with Dunn’s multiple comparisons test was used where **P < 0.01. (F) The frequency of active caspases in miR-155–sufficient and miR-155–deficient plasmablast B cells at day 4.5 post immunisation. At least two independent experiments were carried out. Statistics calculated using nonparametric Kruksal–Wallis with Dunn’s multiple comparisons test where *P < 0.05, **P < 0.01, ****P < 0.0001. Data are representative of at least two independent experiments.
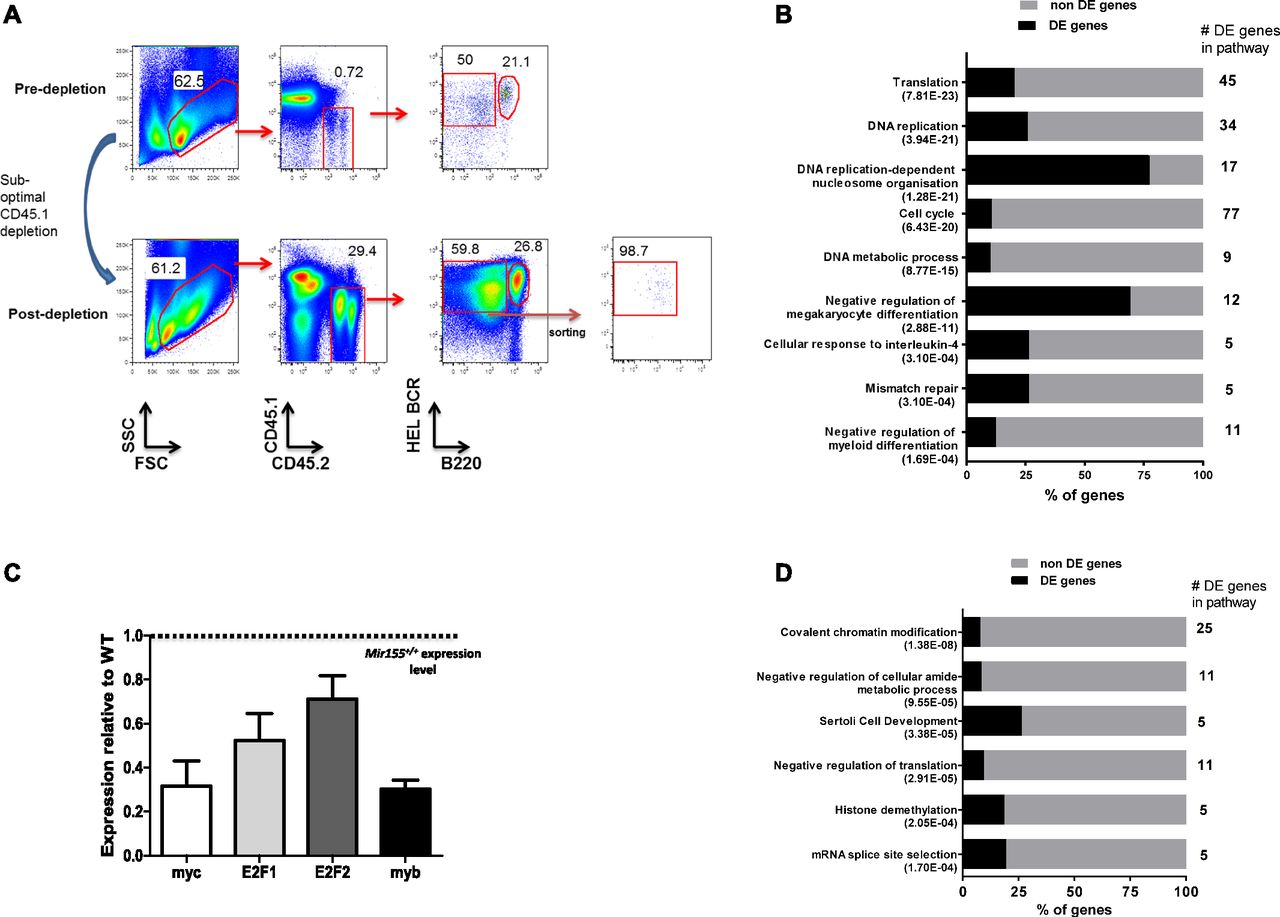
- Figure 4. miR-155 regulates the expression of genes associated with DNA replication.
(A) Representative gating strategy showing CD45.2+ plasmablast B cells before and after suboptimal depletion of CD45.1+ cells. After enrichment, SWHEL Mir155+/+ or Mir155−/− plasmablast B cells were sorted on a FACS Aria to more than 98% purity and analysed by microarray analysis. N = 4–5 Mir155+/+ and 8–10 Mir155−/− mice at day 4.5 post immunisation. (B) Differentially expressed (DE) genes with known functions were discovered by GOrilla analysis using the genes that were down-regulated in SWHEL Mir155−/− plasmablasts compared with SWHEL Mir155+/+ plasmablasts. (C) The mRNA abundance of down-regulated genes with reported roles in DNA replication and function in Mir155−/− plasmablasts relative to Mir155+/+ plasmablasts (dotted line) was confirmed by RT-qPCR. Expression values are normalized to HPRT, determined from three to four biological replicates from three to four independent sorting experiments using 4–5 Mir155+/+ or 8–10 Mir155−/− mice per group. (D) GOrilla analysis of the DE genes that were up-regulated in SWHEL Mir155−/− plasmablast B cells compared with SWHEL Mir155+/+ plasmablast B cells.



Supplementary Materials
Table S3 Primer sequences used for RT-qPCR.




{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- No citing articles found.