



Article Figures & Data
Figures
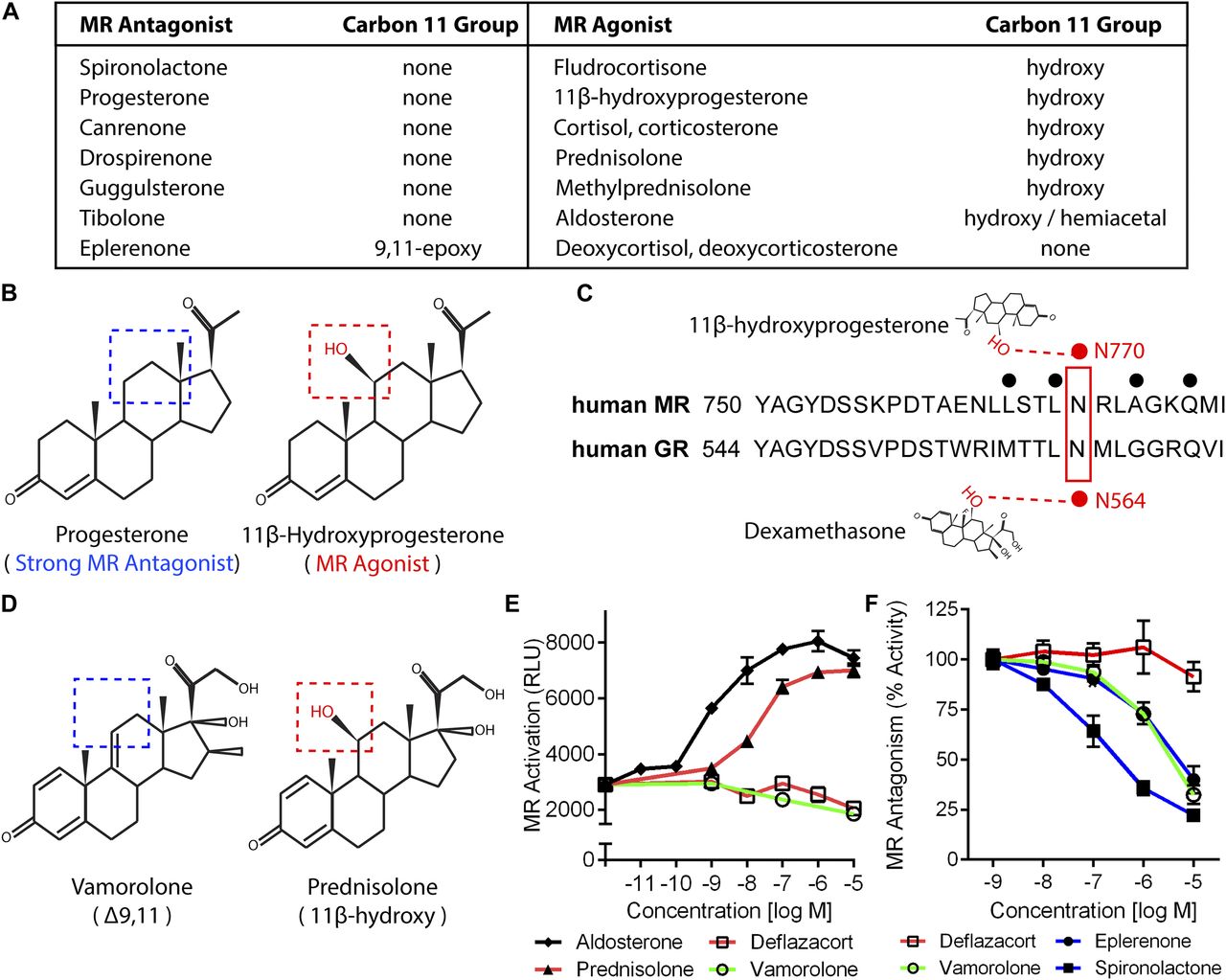
- Figure 1. Vamorolone and MR antagonists lack 11-β hydroxyl groups linked to MR activation.
(A) Table of pharmacological and physiological MR ligands with their carbon 11 group identity provided. (B) Progesterone is a potent MR antagonist, whereas addition of an 11β-hydroxy (11β-Hydroxyprogesterone) results in an agonist compound. (C) The 11β-hydroxy group of hydroxyprogesterone interacts with MR residue N770 via hydrogen bonding. Dexamethasone also interacts with this conserved residue in the GR (N564) via hydrogen bonding. (D) Vamorolone is a Δ9,11 steroid where the 11β position features a carbon–carbon double bond, whereas prednisolone is an 11β-hydroxysteroid. (E) A stable MR reporter cell line was treated with drugs and quantified via chemiluminescence assay to determine their agonist properties. Prednisolone and aldosterone showed MR agonist activity. (F) Reporter cells were treated with drug in combination with a constant E80 dose of aldosterone to determine antagonist properties. Vamorolone acted as an MR antagonist, consistent with eplerenone and spironolactone. (Representative data from three experiments with each dose performed in triplicate; values are mean ± SEM).
- Figure S1. An active metabolite of deflazacort shows no activity in MR reporter assays.
To determine if an active metabolite of deflazacort showed a lack of MR activity consistent with deflazacort, stable MR reporter cells were treated with 21-desacetyl deflazacort, prednisolone, or spironolactone. Used as controls here, prednisolone activated the MR as an agonist, and spironolactone acted as an antagonist to inhibit MR induction by aldosterone. Consistent with deflazacort, we found that the metabolite 21-desacetyl deflazacort avoided MR interaction. (A) Agonist activity was assayed by treating cells with each drug alone in increasing concentrations. (B) Antagonist activity was assayed by pretreating cells with each drug, then adding aldosterone (natural MR agonist) and assaying MR activity 6 h later. (Pred = Prednisolone, 21-Defl = 21-Desacetyl deflazacort, Spiro = Spironolactone).
- Figure 2. CRISPR knockout of the GR shows that anti-inflammatory pathways are GR-dependent.
A frame-shifting deletion mutation was introduced via CRISPR in C2C12 myoblasts to knockout GR expression. (A) The DNA sequence of the GR N-terminus is provided. Base pairs deleted in knockout cells are shown as red, lower case letters. Positions of CRISPR guide RNAs are highlighted in yellow, and genotyping primers in blue. (B) PCR of C2C12 myoblast clones (labeled A through F) identified WT (∼500 bp band) and GR deletion clones (∼300 bp band). (C) Western blot confirmed GR knockout in clone from PCR lane F. (D) Drug-treated wild-type and GR knockout myoblasts were assayed for activation of a GR transactivation target transcript (Gilz). (E) Wild-type and GR knockout myotubes were treated with drug, induced with TNF, and assayed for expression of inflammatory genes regulated by NF-κB (Irf1 and Mcp1). (n = 3–4, *P < 0.05, **P < 0.005, ***P < 0.001, ****P < 0.0001, ANOVA with post hoc versus vehicle [panel D] or + TNF [panel E] control; representative of three experiments; panels [D, E]: [−] = no drug or TNF, [+] = TNF plus vehicle, D = deflazacort, P = prednisolone, V = vamorolone, E = eplerenone).
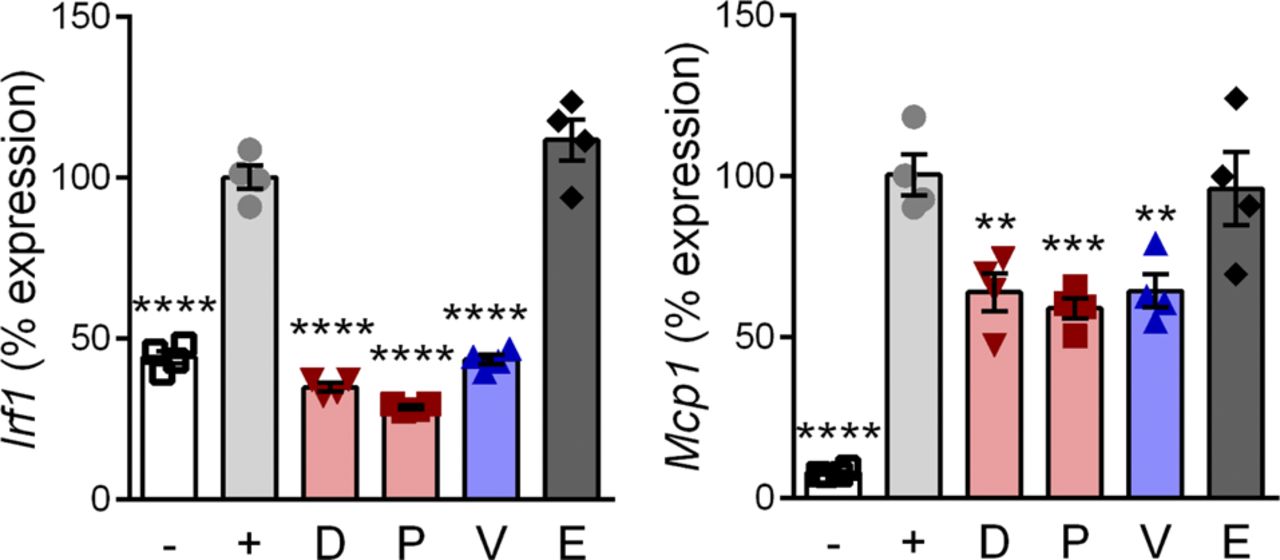
- Figure S2. Vamorolone inhibits inflammatory signaling in macrophage cells.
RAW 264.7 macrophages were pretreated with 10 μM of GR and/or MR ligands and induced with LPS. Expression of NF-κB regulated inflammatory genes (Irf1 and Mcp1) was assayed by qRT-PCR. (n = 4, **P < 0.01, ***P < 0.005, ****P < 0.0001, ANOVA with post hoc versus [+] TNF control in gray; [−] = uninduced, [+] = TNF plus vehicle, D = deflazacort, P = prednisolone, V = vamorolone, E = eplerenone).
- Figure 3. Vamorolone inhibits inflammatory signaling in macrophage and heart cells.
(A) RAW 264.7 macrophages were pretreated with drug at 10 μM and inflammatory signaling was induced for 24 h using LPS. Expression of NF-κB–regulated inflammatory genes (Il1b and Il6) was assayed by qRT-PCR. (B) IL1B and IL6 protein levels were assayed in media from the same experiment via AlphaLISA assay. (C) Primary cardiomyocytes were pretreated with vehicle, vamorolone, or the GR-specific ligand deflazacort, and inflammatory signaling induced with TNF. NF-κB–regulated inflammatory gene expression (Il1b and Il6) was assayed by qRT-PCR. (D) IL6 protein levels were assayed by AlphaLISA. (E) HL-1cells were pretreated with 10 μM drug and induced with TNF for 24 h. Expression of Il6 was assayed by qRT-PCR. (n = 4, **P < 0.005, ****P < 0.0001, ANOVA with post hoc versus [+] TNF control in gray; [−] = no TNF control, [+] = TNF plus vehicle, D = deflazacort, P = prednisolone, V = vamorolone, E = eplerenone).
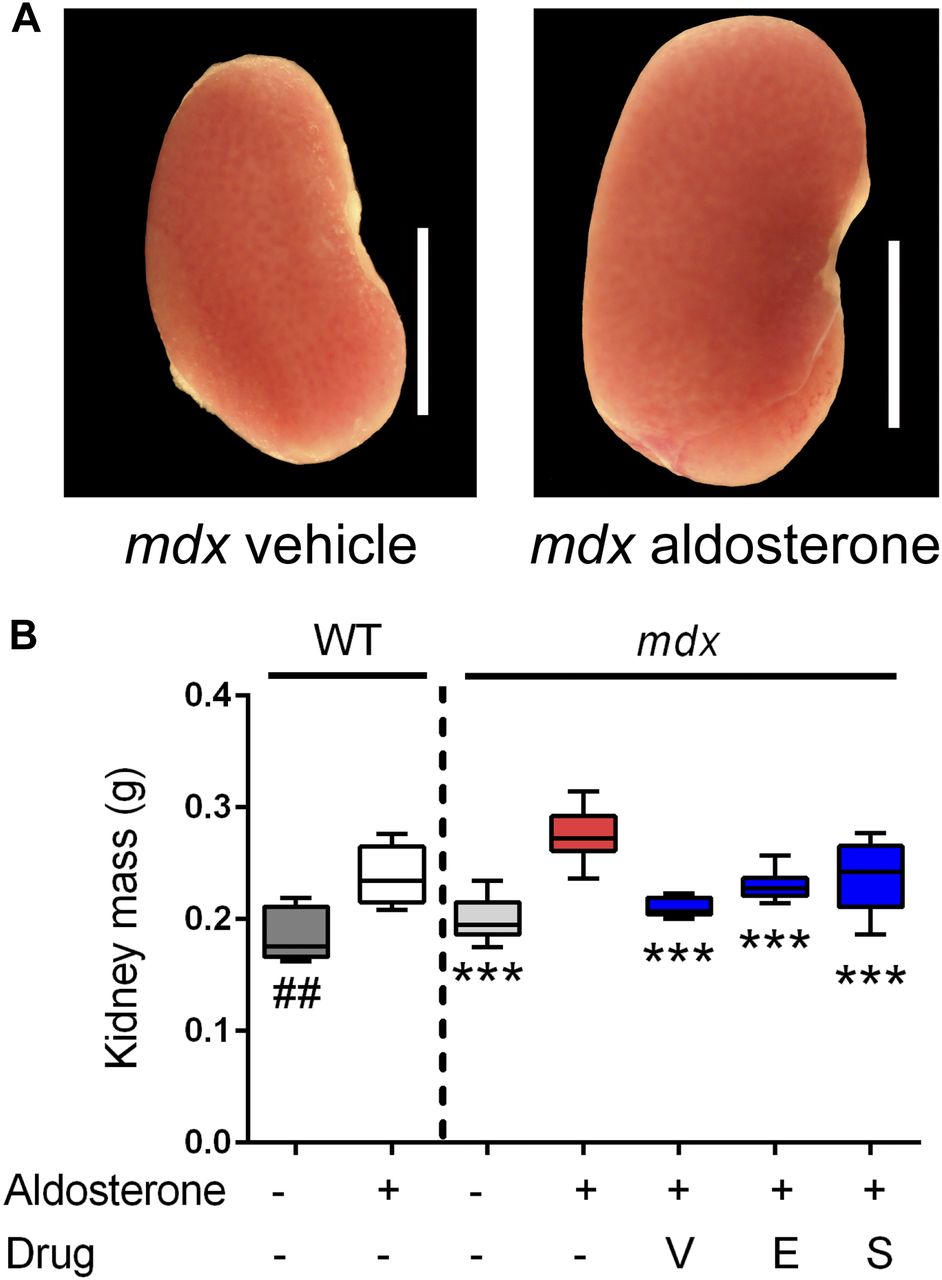
- Figure 4. Vamorolone is an effective MR antagonist in vivo.
Mice were treated with daily oral drug (vehicle, vamorolone, eplerenone, or spironolactone) and implanted with osmotic pumps that secreted either aldosterone or vehicle for 6 wk. Kidney size was assayed as an epithelial MR target tissue, where aldosterone-specificity of the MR is maintained through expression of the glucocorticoid-inactivating enzyme HSD11B2. (A) Representative kidney images are provided. (B) Aldosterone induced an increase in kidney mass in both wild-type and mdx. This was prevented in mdx by all three MR antagonists. (n ≥ 8 per group; ***P < 0.0005, ANOVA with post hoc test versus mdx + aldosterone in red; ##P < 0.005, t test comparing WT groups; V = Vamorolone, E = Eplerenone, S = Spironolactone).
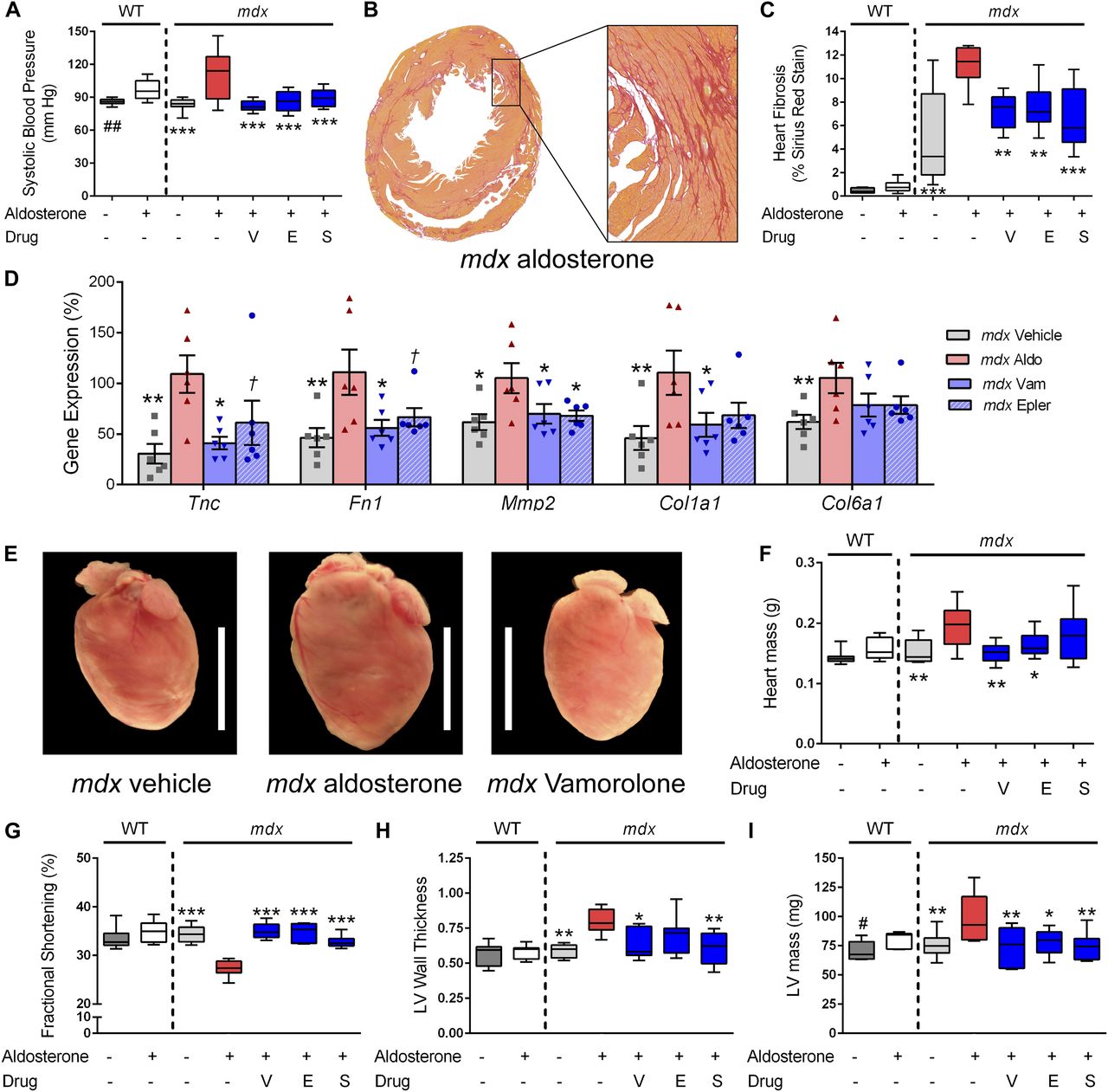
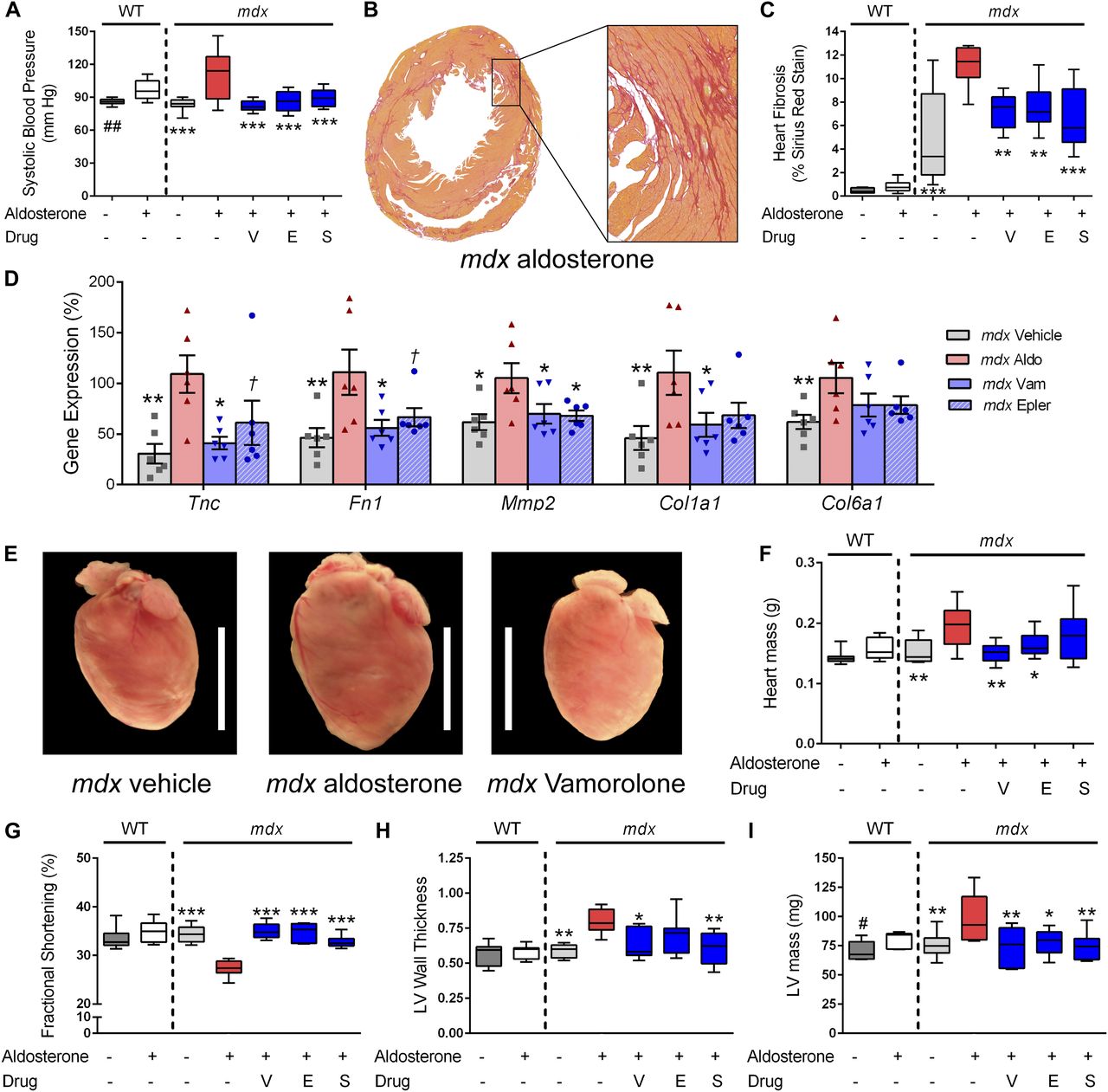
- Figure 5. Vamorolone improves MR-mediated hypertension and mdx cardiomyopathy phenotypes.
Aldosterone introduced via osmotic pumps caused an activation or progression of several phenotypes relevant to cardiomyopathy that were all improved or prevented by vamorolone, eplerenone, and spironolactone. (A) Blood pressure was measured by tail cuff, and increased by aldosterone in both wild-type and mdx mice. (B, C) Sirius Red staining of collagen deposition was used to assay heart fibrosis, where aldosterone increased heart fibrosis specifically in mdx mice in (B) representative images and upon (C) quantification. (D) qRT-PCR of mdx heart tissue was used to assay MR-activated gene expression changes relevant to fibrosis and cardiomyopathy (n = 6 per group). (E, F) Aldosterone increased mdx heart size in (E) representative images and (F) quantitative measurement of heart mass. (G–I) Heart function was assayed by echocardiography, with (G) fractional shortening (H) left ventricular wall thickness (LV wall thickness, in millimetre) and (I) left ventricular mass (LV mass) each showing aldosterone-mediated phenotypes that improved with antagonist treatment. (n ≥ 8; †P < 0.1, *P < 0.05, **P < 0.005, ***P < 0.0005, ANOVA with post hoc versus mdx + aldosterone in red; #P < 0.05, ##P < 0.005, t test comparing WT groups; V = Vamorolone, E = Eplerenone, S = Spironolactone).
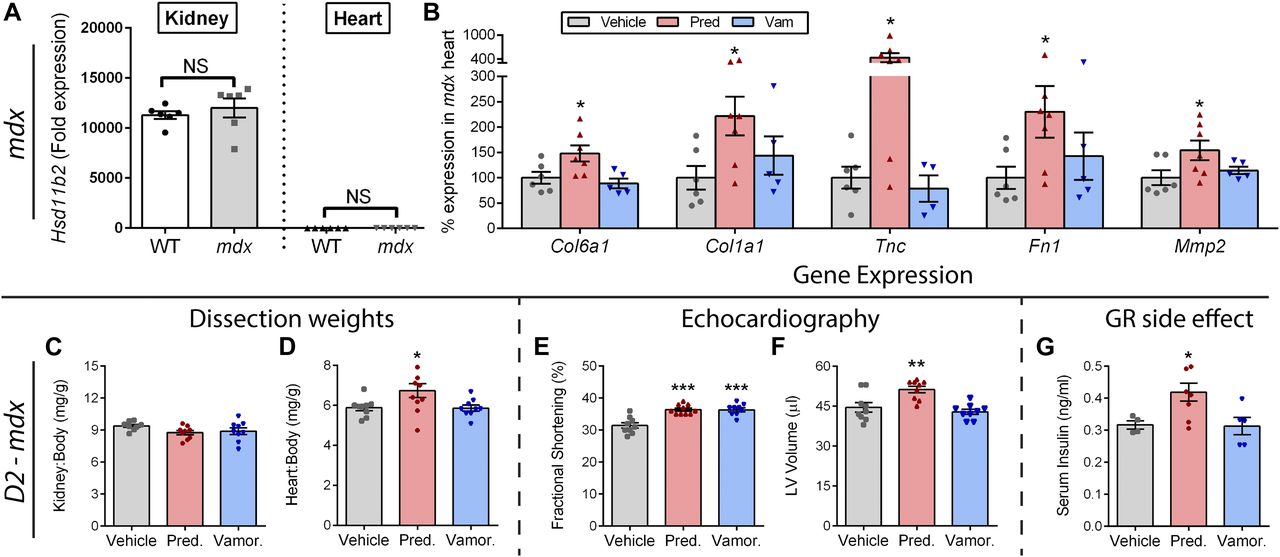
- Figure 6. Prednisolone activates MR and GR side effect pathways in mdx and severe D2-mdx mice.
(A) Expression of the glucocorticoid-metabolizing gene Hsd11b2 was assayed by qRT-PCR in kidneys and hearts from wild-type and mdx mice. Lack of substantial expression in the heart indicated that the heart is sensitive to MR activation by prednisolone. (B) Standard mdx mice on a BL10 background were treated with vehicle, prednisolone (5 mg/kg), or vamorolone (45 mg/kg) for 4 months, beginning at 6 weeks of age. The heart was assayed for expression of MR-regulated genes via qRT-PCR and expressed as % vehicle levels (n ≥ 5). (C–G) Severe D2-mdx mice on an LTBP4 modifier background were treated for 8 weeks with daily oral vehicle, vamorolone (30 mg/kg), or prednisolone (5 mg/kg), beginning at 2 mo of age (n ≥ 9). (C, D) Kidney and heart mass at trial conclusion. Note, kidney expresses the glucocorticoid-inactivating enzyme (HSD11B2) that protects it from prednisolone, whereas the heart does not. Prednisolone increased D2-mdx heart size but not kidney size. (E, F) Echocardiography was performed near the trial endpoint. (E) Fractional shortening increased with both vamorolone and prednisolone. (F) Left ventricular volume (end diastolic) did not change with vamorolone but increased with prednisolone, consistent with premature dilatation. (G) Serum insulin levels increased with prednisolone (n ≥ 5), consistent with GR-mediated, metabolic side effects of glucocorticoids. (*P < 0.05, **P < 0.005, ***P < 0.0005; one panel B outlier removed after significant Grubbs test; (A) t test of mdx versus wild-type for each tissue, (B–G) ANOVA with post hoc versus vehicle in gray).
- Figure 7. Working model of dual-receptor drug mechanisms in epithelial and non-epithelial tissues.
Left Panel: Epithelial MR target tissues (e.g., kidney, colon, skin) co-express the HSD11B2 enzyme to prevent over-activation of the MR by glucocorticoids. This protein metabolizes 11β-hydroxysteroids (cortisol and prednisolone) into their inactive ketone states (cortisone and prednisone) within that tissue. Aldosterone is not metabolized by HSD11B2, thus enabling aldosterone-specific signaling in vivo. We find that aldosterone challenge increases kidney size, whereas prednisolone treatment does not, consistent with this model. Right Panel: Non-epithelial MR target tissues express the MR but do not express the HSD11B2 enzyme (heart, brain, skin, fat, immune cells). These tissues may have increased risk of damage from MR activation by glucocorticoids. The heart is particularly important here, as (1) DMD patients naturally develop cardiomyopathy, and (2) a “second hit” provided by the DMD disease could worsen MR-mediated heart pathologies (such a “second hit” is seen in other aldosterone models). We find that both aldosterone and prednisolone increase mdx heart size and fibrosis, consistent with increased MR activation. Both Panels: Vamorolone is a potent MR antagonist that protects both epithelial and non-epithelial tissue types. (Black = physiological ligands, Blue = Vamorolone, Red = Prednisolone; Aldo = Aldosterone, Cort = Cortisol, HSD11B2 = 11β-hydroxysteroid dehydrogenase, Pred = Prednisolone, Vam = Vamorolone).








Tables
- Table 1.
Summary of drug sub-activity profiles on three promoter types for GR and MR ligands.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
