



Article Figures & Data
Figures
- Figure 1. REC114 is essential for spermatogenesis and oogenesis.
(A) Conserved domains and organization of REC114. The conserved motifs are SSM1 to 7 (Kumar et al, 2010). Secondary structures were predicted with PSIPRED v3.3 (http://bioinf.cs.ucl.ac.uk/psipred/). Pink cylinders, α−helices; yellow arrows, β−sheets. Exons (1 to 6) are shown as dark and light blue rectangles. (B) REC114 is not detected in Rec114−/− mice. Western blot (WB) analysis of total testis extracts from WT, Mei4−/−, and Rec114−/− as control, prepubertal mice (14 d post-partum, dpp) with anti-REC114 (central panel), with anti-IHO1 (right panel), and anti-REC114 antibodies after immunoprecipitation of REC114 (left panel). (C) Spermatogenesis is defective in Rec114−/− mice. Periodic acid-Schiff staining of testis sections from 9-wk-old Rec114+/+ and Rec114−/− mice. (D) Oogenesis is defective in Rec114−/− mice. Hematoxylin and eosin staining of ovary sections from 2-wk-old Rec114+/+ and Rec114−/− mice. (E) Quantification of primordial, primary, and secondary follicles in ovaries from 2-wk-old and 8-wk-old Rec114+/+, Rec114+/−, and Rec114−/− mice. At 2 wk of age, the numbers (mean ± SD) of primordial (blue circles), primary (green circles), and secondary (red circles) follicles were 32.1 ± 17.9, 8.4 ± 4.1, and 23.1 ± 6.5, respectively, for Rec114+/+ (n = 3) and Rec114+/− (n = 1) mice (Rec114+/+ and Rec114+/− data were pooled, n sections = 21 in total) and 0.6 ± 0.9, 2.3 ± 1.8, and 16.1 ± 6.8, respectively, for Rec114−/− mice (n = 5; n sections = 21). At 8 wk of age, the numbers (mean ± SD) of primordial (blue circles), primary (green circles), and secondary (red circles) follicles were 6.3 ± 5.0, 4 ± 1.8, and 7.2 ± 3.8, respectively, for Rec114+/+ mice (n = 1; n sections = 6), and 0.1 ± 0.3, 0 ± 0, and 0.5 ± 1.0, respectively, for Rec114−/− mice (n = 2; n sections = 10). P values were calculated with the Mann–Whitney two-tailed test. Sz, spermatozoa; St, round spermatid; Sp, spermatocyte.
- Figure S1.
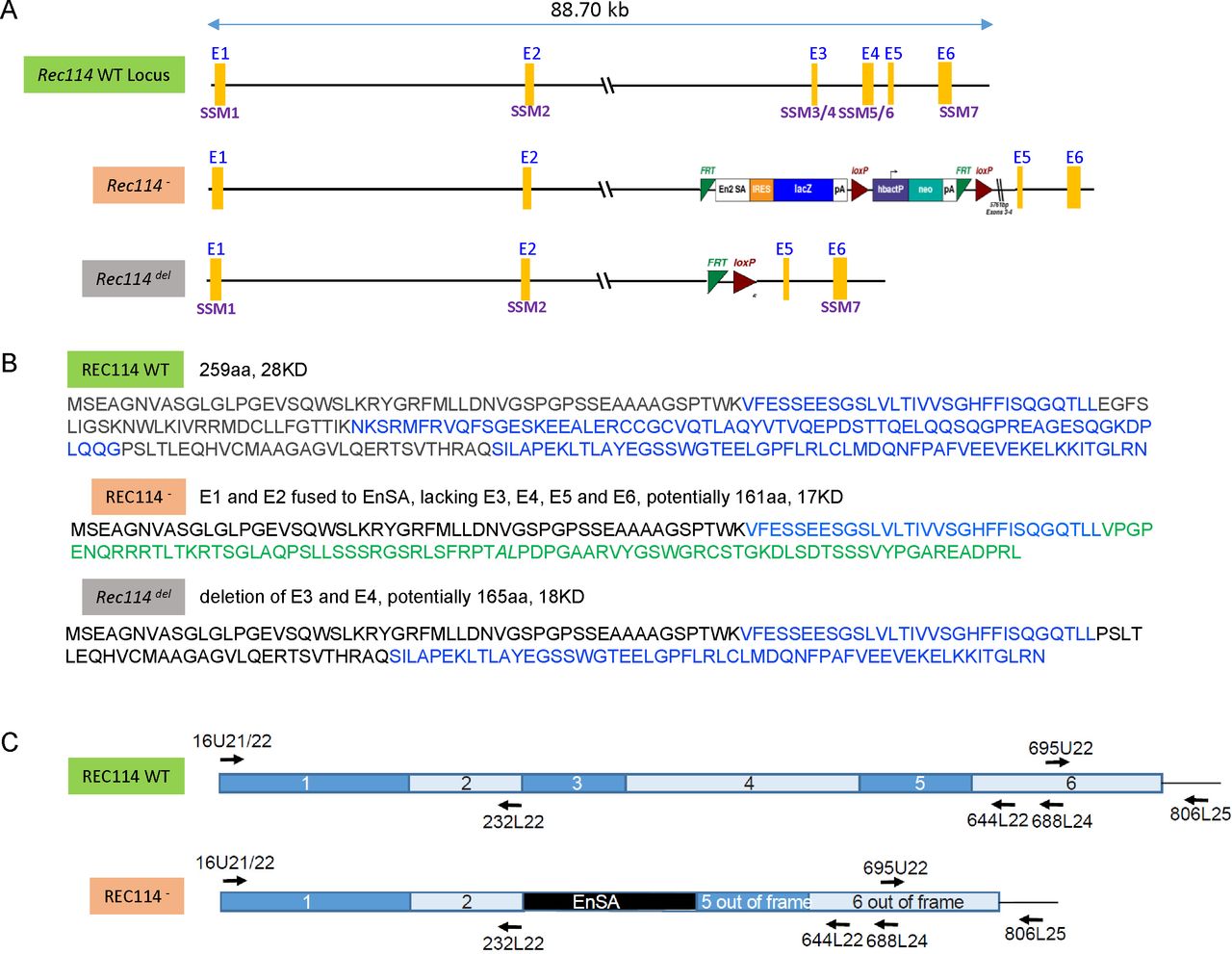
(A) Map of the genomic region including the WT Rec114, Rec114−, and Rec114del alleles. The six Rec114 exons are labelled E1 to E6. The conserved motifs are labelled SSM1 to SSM7 (Kumar et al, 2010). Note that SSM2 position was revised by Tesse et al (2017). The Rec114del allele was obtained by expression of the Flip recombinase (Flp) in mice carrying the Rec114− allele. (B) Proteins potentially encoded by the different Rec114 alleles. (C) Schematic representations of the WT and Rec114− cDNAs. The positions of oligonucleotides used for RT-PCR are indicated on the cDNA maps. DNA sequencing of cDNAs from Rec114−/− mice revealed after exon 2 an insertion of a 113b DNA sequence from the cassette which contains a splice acceptor site (EnSA). Exon 5 and 6 are out of frame.
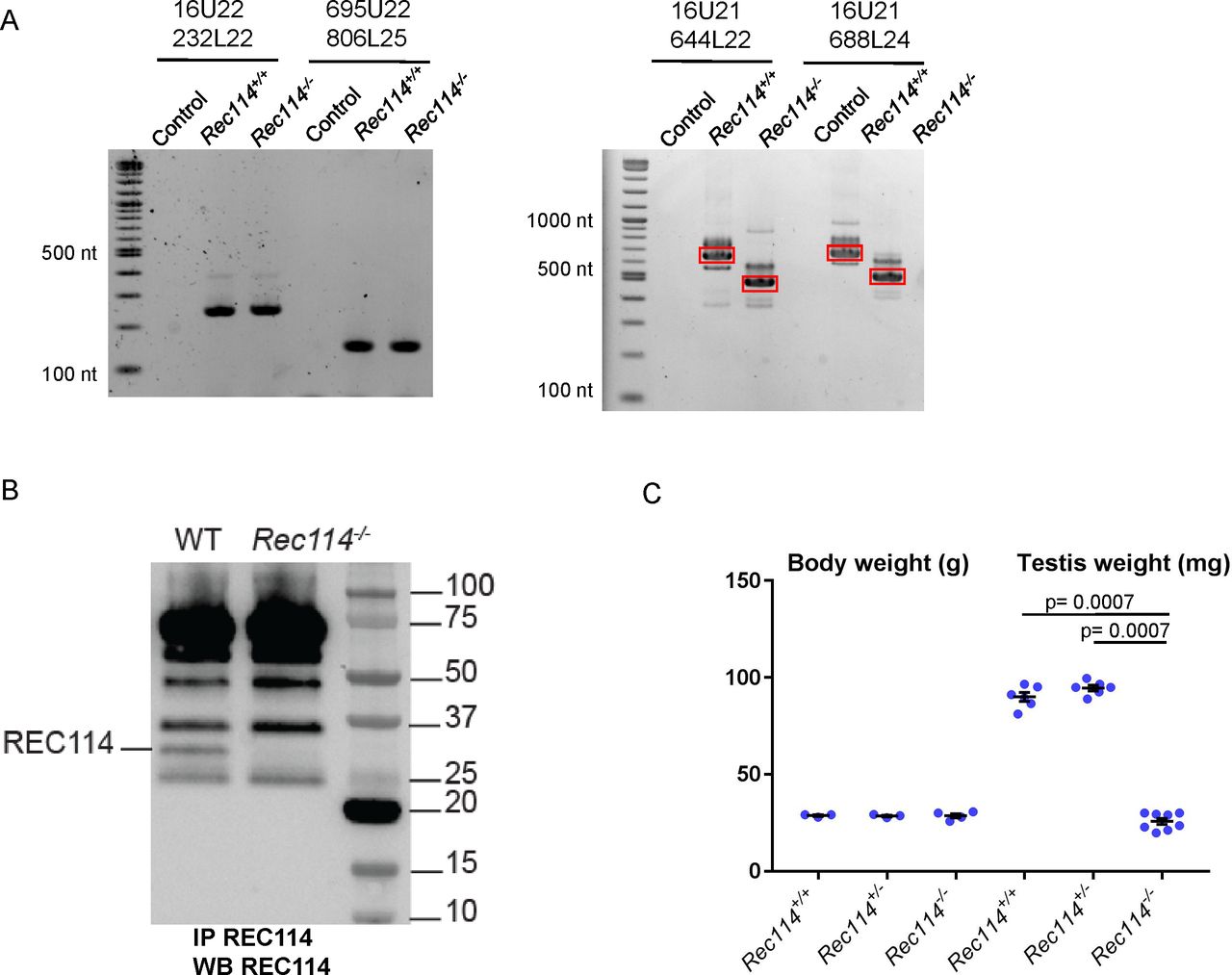
- Figure S2.
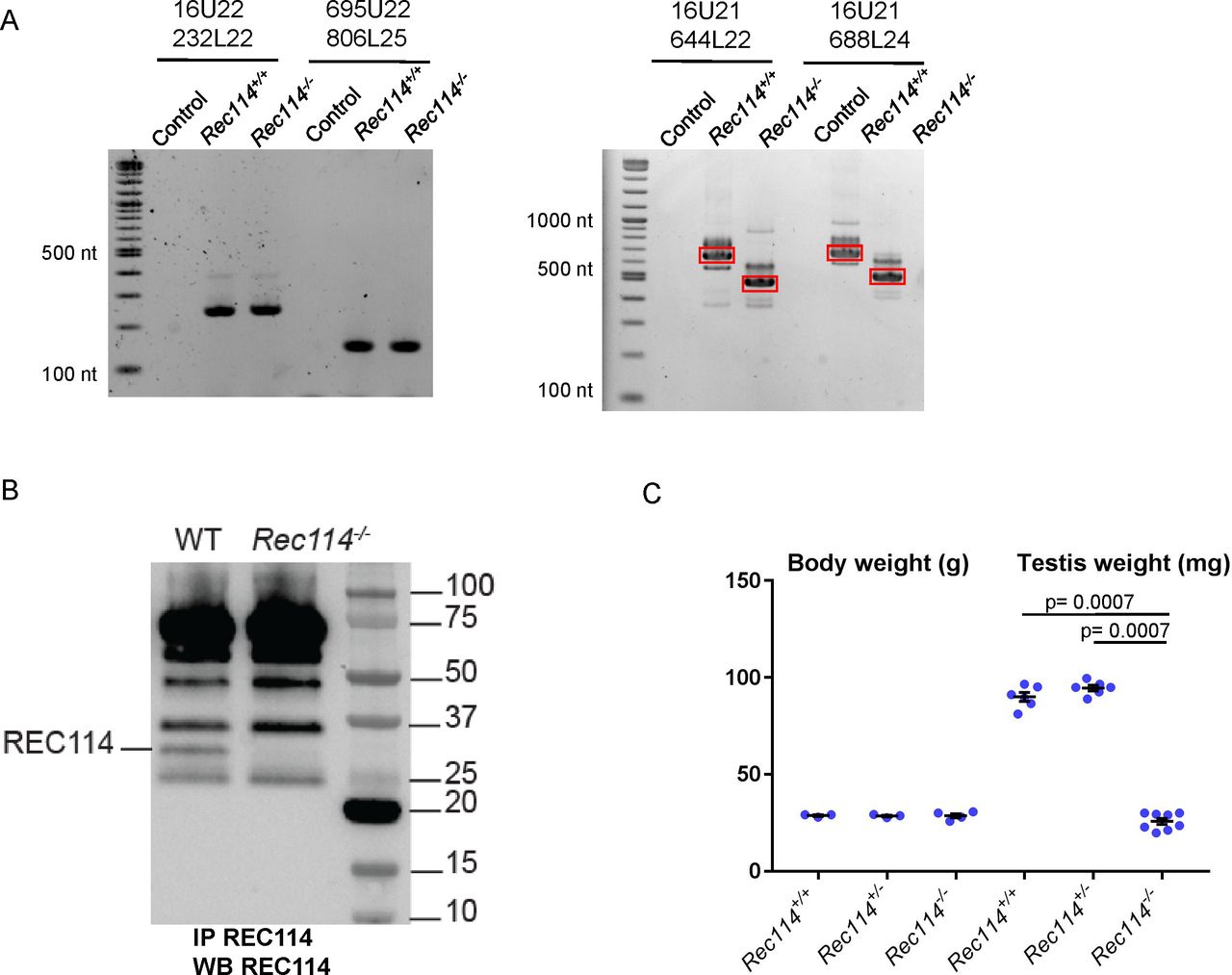
(A) RT-PCR analysis on RNA extracted from testes of Rec114+/+ and Rec114−/− 14 dpp mice. Controls are PCR without cDNA. The RT-PCR products were separated by agarose gel electrophoresis. Purified PCR products that were sequenced are highlighted in red rectangles. With primer pairs 16U21/644L22 and 16U21/688L24, the PCR products obtained from Rec114−/− mice migrate faster compared with Rec114+/+ mice because of the deletion of DNA sequences from exon 3 and 4 and insertion of a 113b DNA sequence from the cassette (EnSA). (B) Western blot analysis of total testis extracts from WT, 13 dpp and Rec114−/−, 12 dpp after immunoprecipitation with anti-REC114 antibodies, and detection with the same antibody. (C) Testis weight is reduced in Rec114−/− mice compared with Rec114+/+ and Rec114+/− mice. Body and testis weights (mean ± SD) were measured in 8- to 10-wk-old Rec114+/+ (testis weight 90 ± 5.7; n = 6), Rec114+/− (testis weight 94.6 ± 3.6 mg; n = 6), and Rec114−/− (testis weight 25.8 ± 4.3 mg; n = 6) males. P values were calculated with the Mann–Whitney two-tailed test.
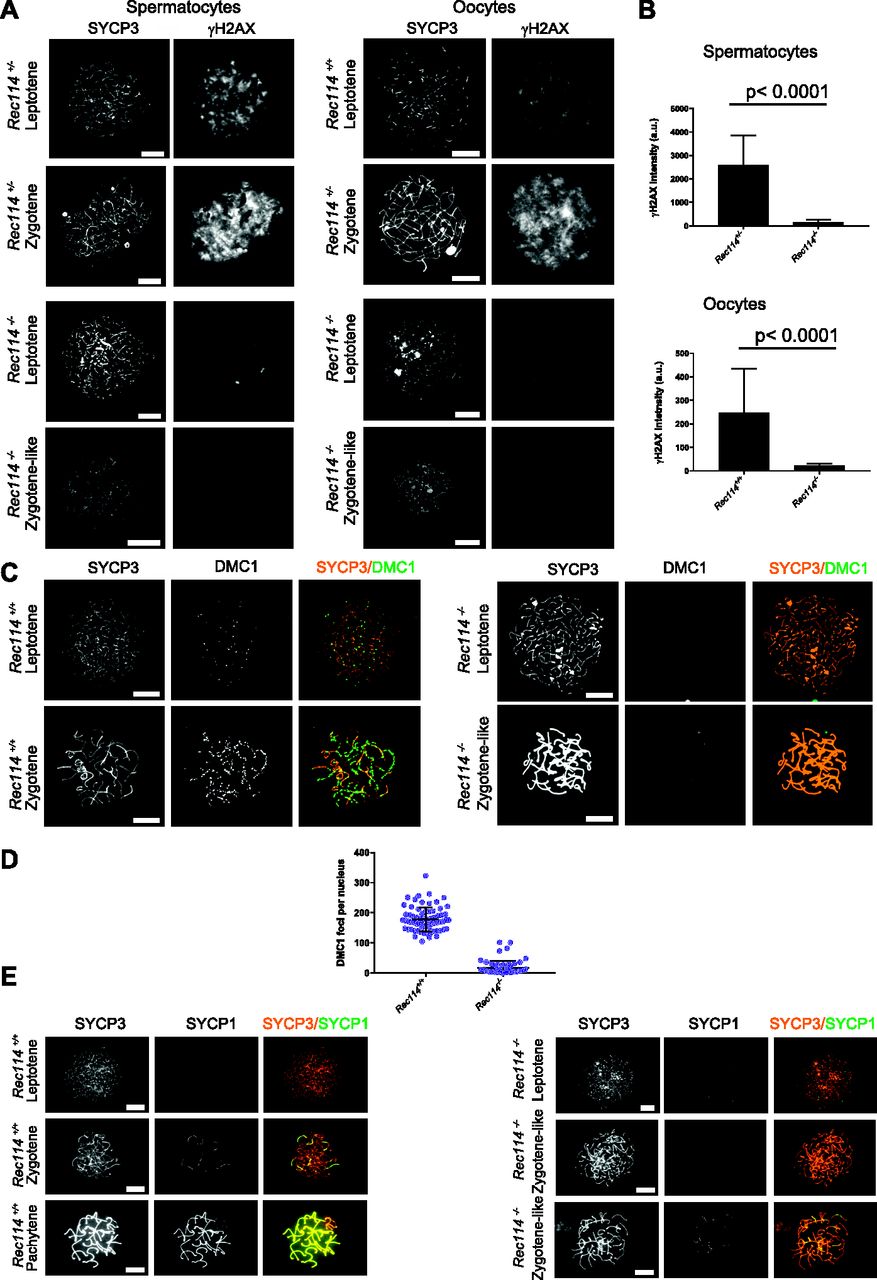
- Figure 2. Rec114−/− mice show defects in DSB formation and homologous synapsis.
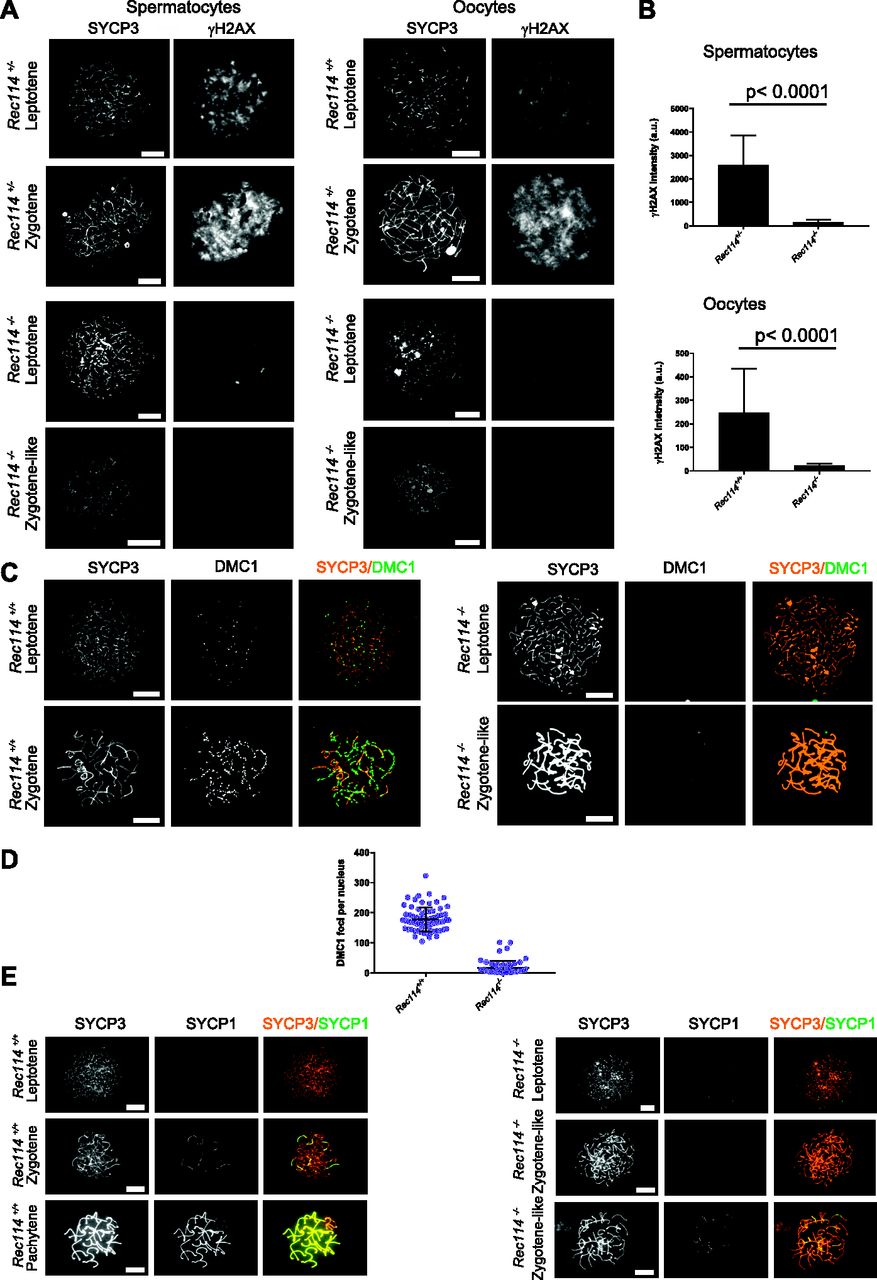
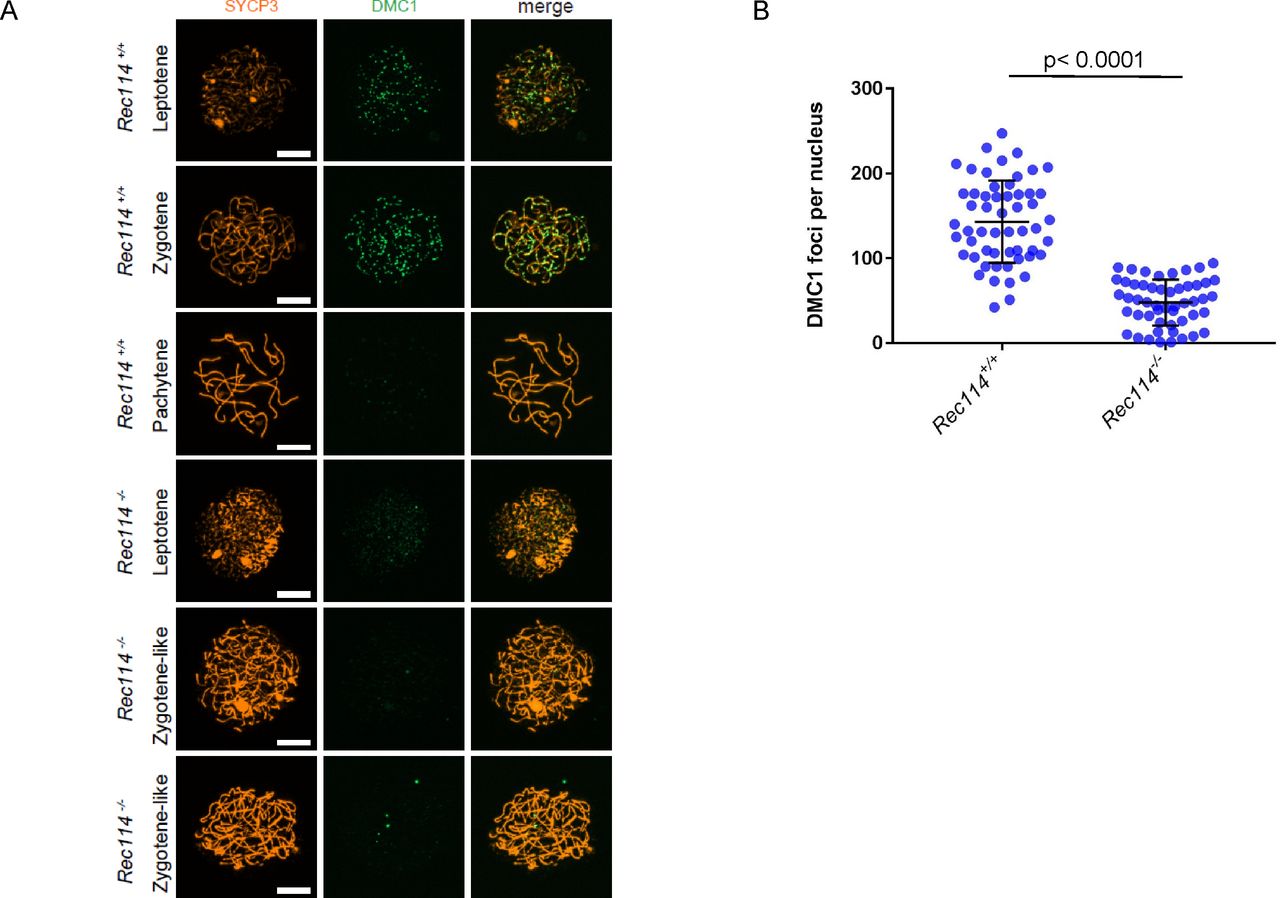
(Α) Immunostaining of γH2AX and SYCP3 in spermatocytes from 13 dpp Rec114+/− and Rec114−/− males, and from E15 (15 d of embryonic development) Rec114+/+, Rec114+/−, and Rec114−/− oocytes. In Rec114−/− spermatocytes and oocytes, no pachynema could be observed and spermatocytes or oocytes with partially synapsed chromosomes were defined as zygotene-like. Scale bar, 10 μm. (B) Quantification of the total γH2AX signal per nucleus (mean ± SD; a.u, arbitrary units) on spreads from leptotene spermatocytes (13 dpp) and from leptotene oocytes (E15): 2,597 ± 1,261 and 165 ± 95 in Rec114+/− and Rec114−/− males, respectively (n = 53 and 50); 248 ± 187 and 23 ± 7 in Rec114+/+ and Rec114−/− females, respectively (n = 48 and 47). P values were calculated with the Mann–Whitney two-tailed test. (C) Immunostaining of DMC1 and SYCP3 in spermatocytes from 15 dpp Rec114+/+ and Rec114−/− males. Scale bar, 10 μm. (D) Quantification of DMC1 foci (mean ± SD) in leptotene and zygotene spermatocytes from Rec114+/+ and Rec114−/− mice (178.6 ± 39.9 and 17.2 ± 23.0 in Rec114+/+ and Rec114−/− males, respectively; n = 71 and 59). P < 0.0001 (Mann–Whitney two-tailed test). (E) Immunostaining of SYCP1 and SYCP3 in spermatocytes from 15 dpp Rec114+/+ and Rec114−/− mice. Scale bar, 10 μm.
- Figure S3. Reduction of DMC1 foci in Rec114−/− oocytes
(A) Immunostaining of DMC1 and SYCP3 in oocytes from E15 Rec114+/+ and Rec114−/− females. Scale bar, 10 μm. (B) Quantification of DMC1 foci (mean ± SD) in leptotene and zygotene oocytes from E15 Rec114+/+ and Rec114−/− E15 mice (143.0 ± 48.7 and 47.8 ± 27.1, respectively; n = 55 and 51 nuclei). P value was calculated with the Mann–Whitney two-tailed test.
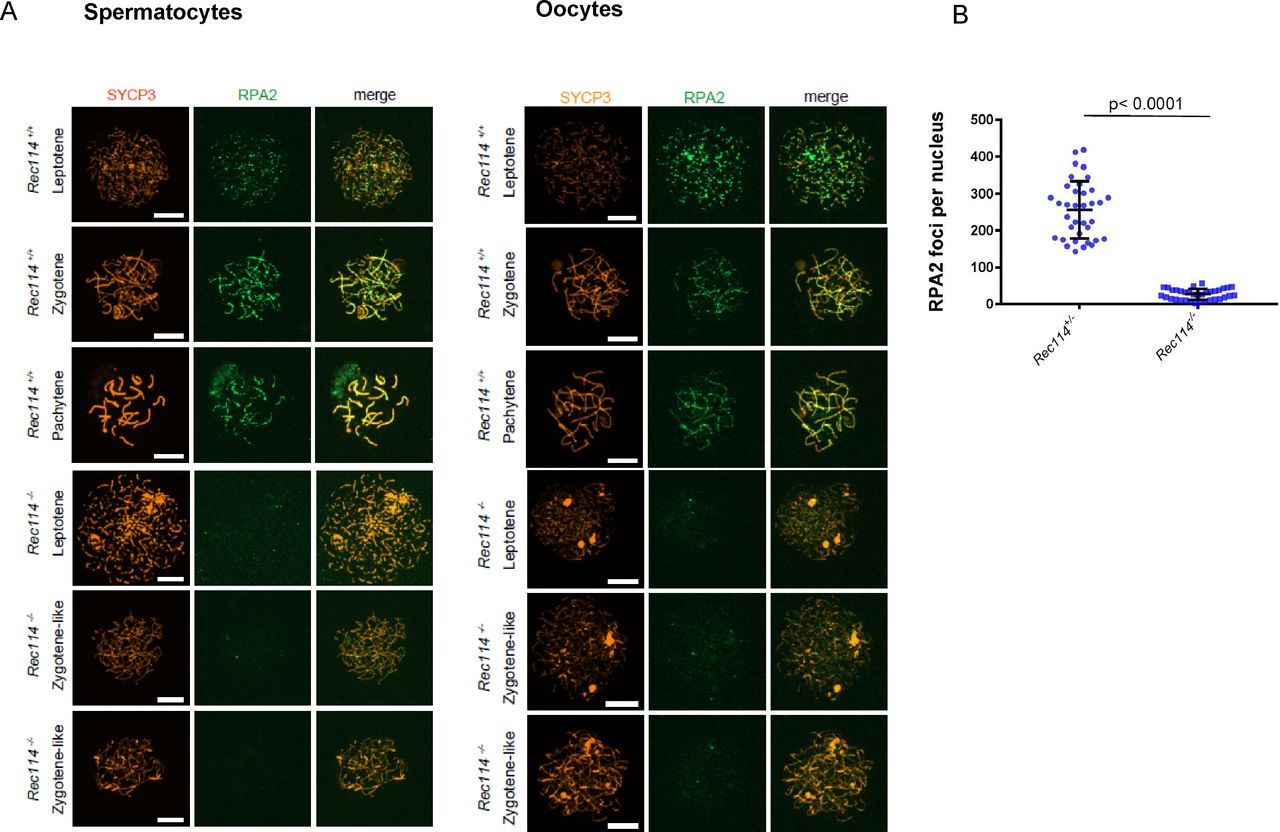
- Figure S4. Reduction of RPA2 foci in Rec114−/− meiocytes
(A) Immunostaining of replication protein A2 (RPA2) and SYCP3 in 15 dpp spermatocytes and E15 oocytes from Rec114+/+ and Rec114−/− mice. Scale bar, 10 μm. (B) Quantification of RPA2 foci (mean ± SD) in leptotene and zygotene oocytes from E15 Rec114+/− and Rec114−/− E15 mice (255.9 ± 77.7 and 26.5 ± 14.8, respectively; n = 36 and 35 nuclei). P value was calculated with the Mann–Whitney two-tailed test.
- Figure S5. Reduction of DSB repair foci is not due to absence of SPO11 in Rec114−/− spermatocytes
(A) Immunostaining of RAD51 and SYCP3 in spermatocytes from 15 dpp Rec114+/+ and Rec114−/− mice. Scale bar, 10 μm. (B) Detection of SPO11 in Rec114−/− testis extracts. Immunoprecipitation of SPO11 in testis extracts from WT, Rec114−/−, Spo11−/−, and Spo11YF/YF adult mice. Only the SPO11β variant (44KD) is detected in these conditions.
- Figure S6. Deficiency in SYCP1 assembly in Rec114−/− oocytes
Immunostaining of SYCP1 and SYCP3 in oocytes from Rec114+/+ females (E15, E15, and E16 for leptotene, zygotene, and pachytene, respectively) and Rec114−/− females (E15, E16, and E17 for leptotene, zygotene-like, and zygotene-like, respectively). Scale bar, 10 μm.
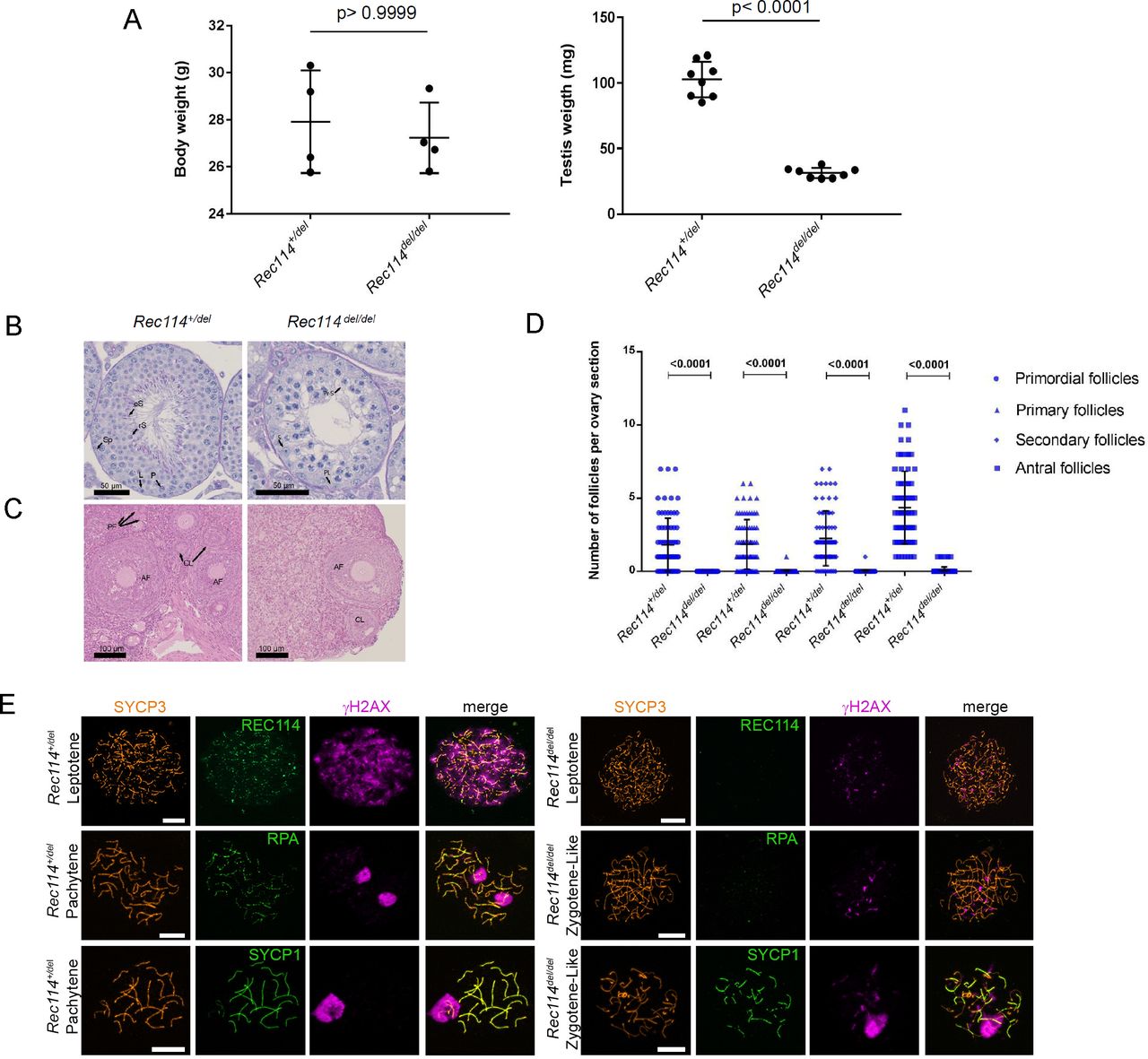
- Figure S7. The Rec114del allele confers the same phenotypes as the Rec114− allele
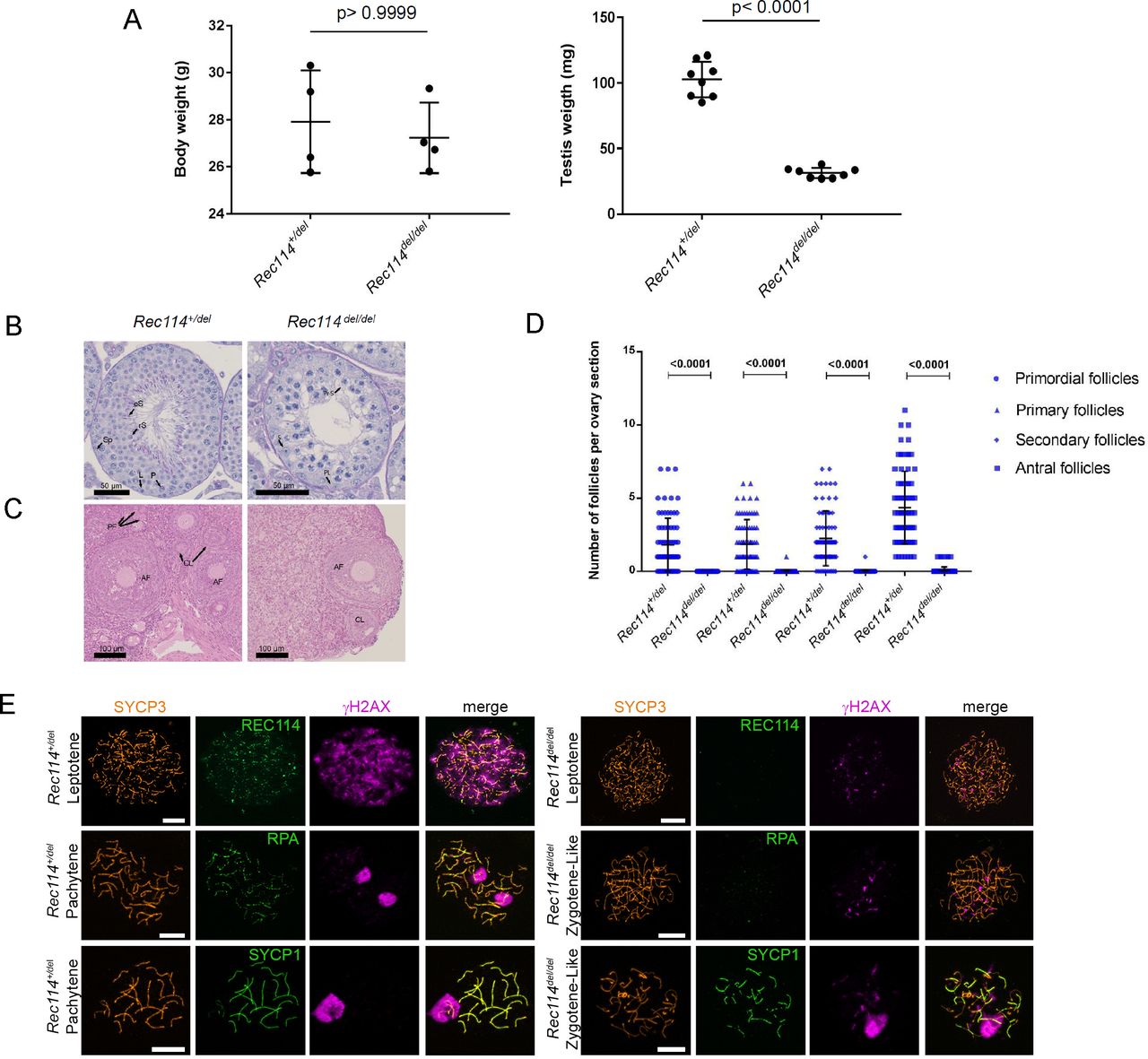
(A) Testis weight is reduced in Rec114del/del mice compared with Rec114+/del mice. Body and testes weights (mean ± SD) were measured in 8- to 10-wk-old Rec114+/del (testis weight 102.7 ± 13.6 mg; n testes = 8) and Rec114del/del mice (testis weight 31.3 ± 3.9 mg; n testes = 8). P values were calculated with the two-tailed Mann–Whitney test. (B) Periodic acid-Schiff staining of testes sections from 9-wk-old Rec114+/del and Rec114del/del mice. (C) Hematoxylin and eosin staining of ovary sections from 9-wk-old, Rec114+/del and Rec114del/del mice. (D) Quantification of primordial, primary, secondary and antral follicles in ovaries from 9-wk-old Rec114+/del and Rec114del/del mice. P values were calculated with the Mann–Whitney two-tailed test. (E) Immunostaining of SYCP3, REC114, and γH2AX in leptotene, of SYCP3, RPA, and γH2AX in zygotene, and of SYCP3, SYCP1, and γH2AX in zygotene-like or pachytene spermatocytes from 9-wk-old Rec114del/del and Rec114+/del mice. Scale bar, 10 μm. AF, antral follicle; CL, corpus luteus; eS, elongated spermatid; L, leptotene; P, pachytene; PF, primary follicle; PL, pre-leptotene; Pr-S, primary spermatocyte; rS, round spermatid; S, Sertoli cell; Sp, spermatogonia.
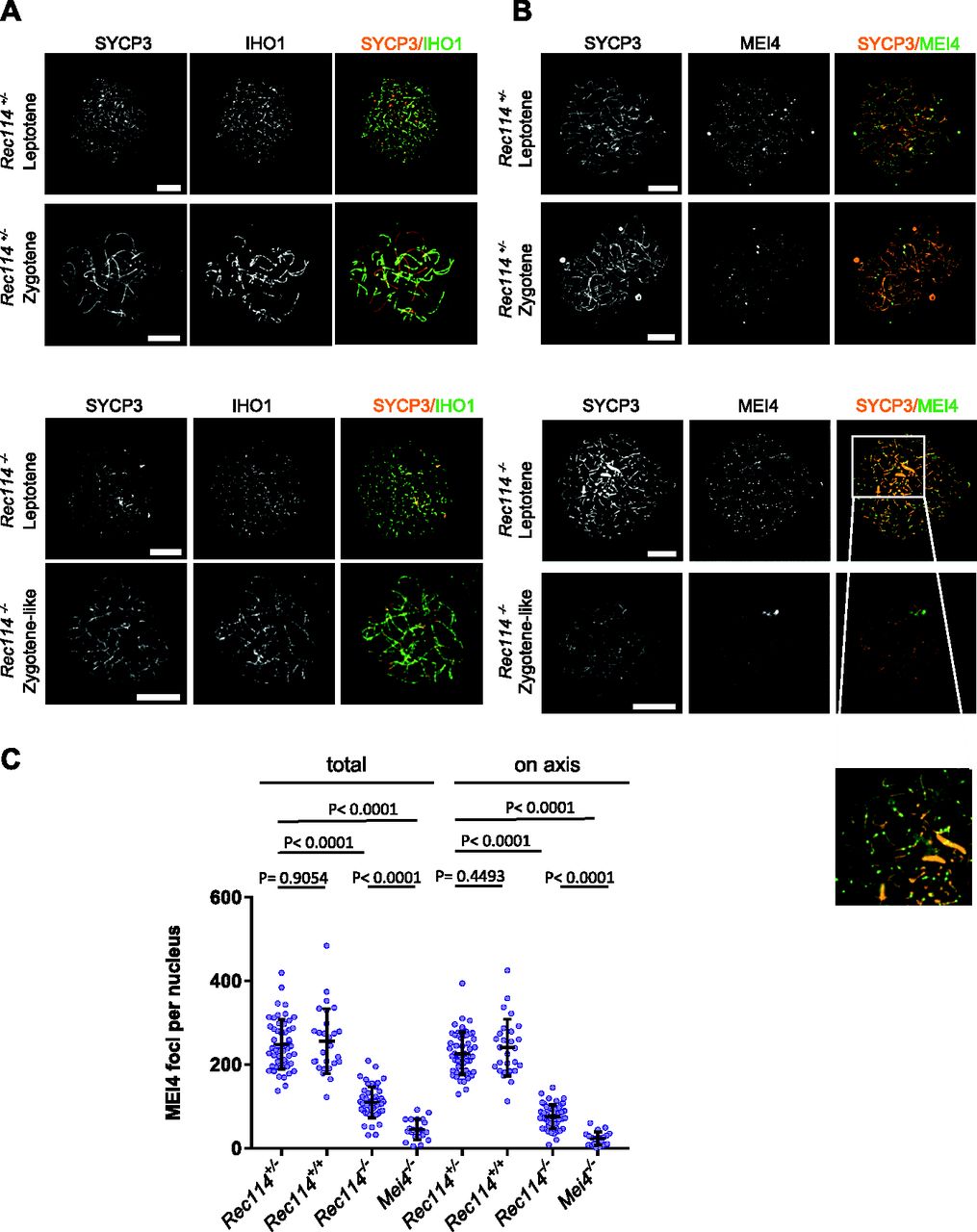
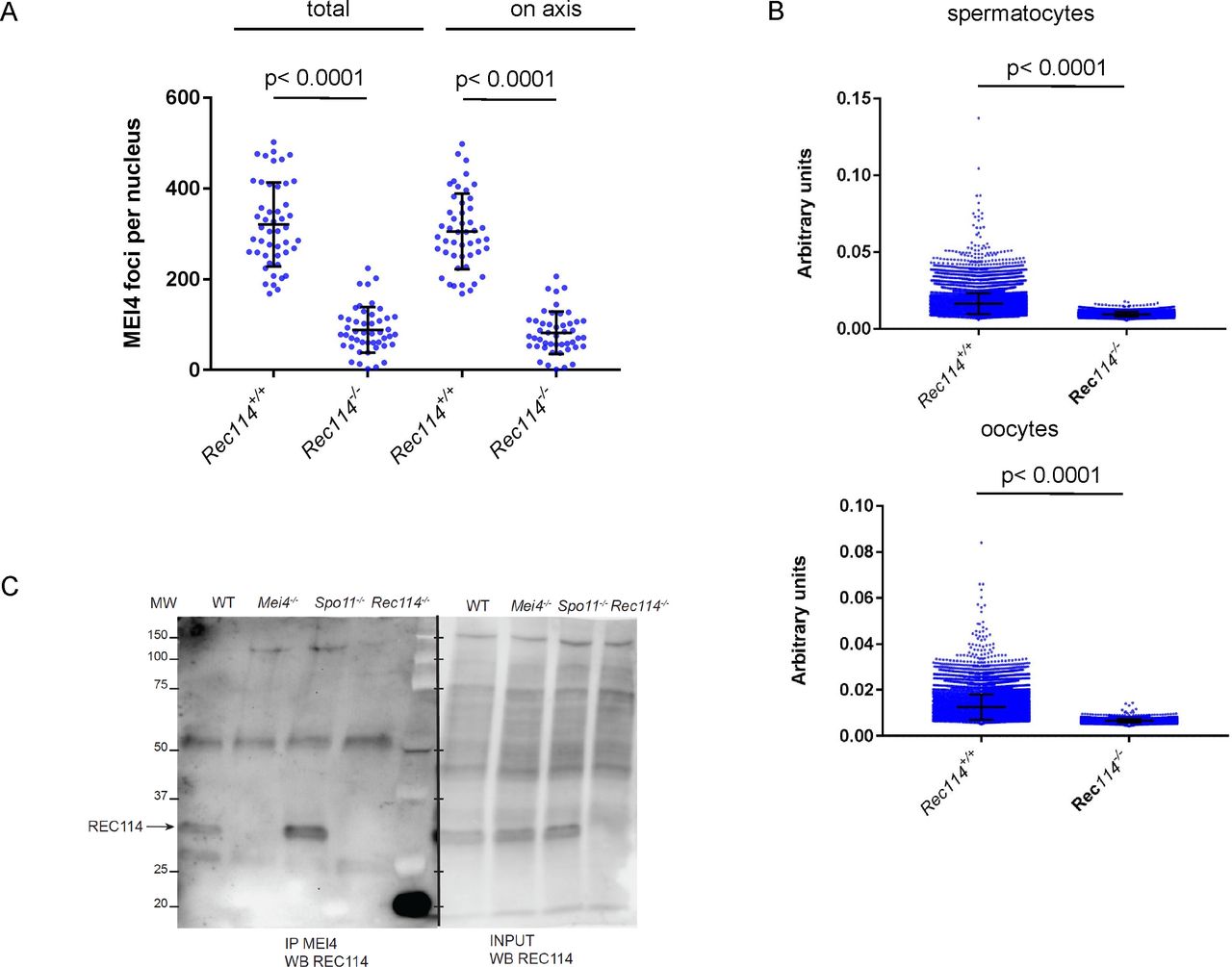
- Figure 3. REC114 is required for robust MEI4 foci localization.
(A) Immunostaining of IHO1 and SYCP3 in early prophase spermatocytes from 13 dpp Rec114+/− and Rec114−/− males. Scale bar, 10 μm. (B) Immunostaining of MEI4 and SYCP3 in early prophase spermatocytes from 13 dpp Rec114+/− and Rec114−/− males. Scale bar, 10 μm. (C) Quantification of MEI4 foci in leptotene spermatocytes from 13 dpp Rec114+/−, Rec114+/+, Rec114−/−, and Mei4−/− males. The numbers (mean ± SD) of total foci and of foci on chromosome axes (co-localized with SYCP3) were: 248 ± 59 and 226 ± 51 for Rec114+/− (n = 53 nuclei), 256 ± 78 and 241 ± 68 for Rec114+/+ (n = 27), 109 ± 37 and 76 ± 29 for Rec114−/− (n = 50), 45 ± 25, and 23 ± 16 for Mei4−/− (n = 20), respectively. P values were calculated with the Mann–Whitney two-tailed test.
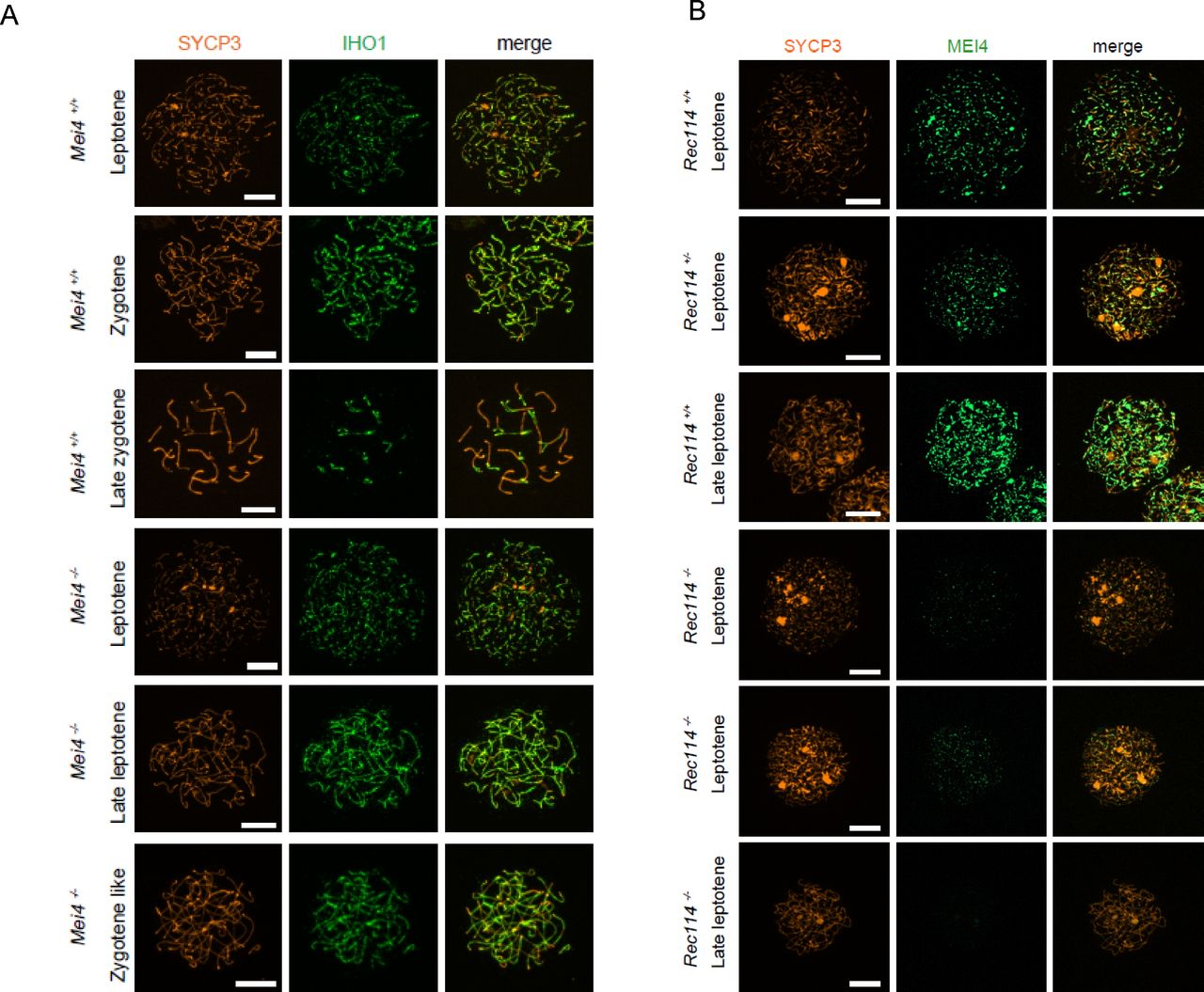
- Figure S8. IHO1 localisation does not require MEI4, and MEI4 localisation requires REC114
(A) Immunostaining of IHO1 and SYCP3 in early prophase spermatocytes from 13 dpp Mei4+/+ and 12 dpp Mei4−/− males. Scale bar, 10 μm. (B) Immunostaining of MEI4 and SYCP3 in early prophase oocytes from E15 Rec114+/+, Rec114+/−, and Rec114−/− females. Scale bar, 10 μm.
- Figure S9. Quantification of MEI4 foci and co-immunoprecipitation with REC114
(A) Quantification of MEI4 foci (mean ± SD) in leptotene oocytes from E15 Rec114+/+ and Rec114−/− females. The numbers of total foci and foci on chromosome axis (co-localized with SYCP3) were 321 ± 93 and 305 ± 83 in in Rec114+/+ and 88 ± 50 and 82 ± 47 in Rec114−/− oocytes; n = 48 and 47). P values were calculated with the Mann–Whitney two-tailed test. (B) Quantification of intensity (arbitrary units) of axis-associated MEI4 foci (mean ± SD) in leptotene spermatocytes and oocytes from Rec114+/+ and Rec114−/− mice: 0.0164 ± 0.0068 and 0.0094 ± 0.0015 in Rec114+/+ and Rec114−/− spermatocytes (n = 12,001 and 3,775, respectively) and 0.0125 ± 0.0054 and 0.0066 ± 0.0009 in Rec114+/+ and Rec114−/− oocytes (n = 14,661 and 3,853, respectively). P values were calculated with the Mann–Whitney two-tailed test. (C) Detection of REC114 after MEI4 immunoprecipitation. Total testis extracts from 14 dpp Rec114+/+ (WT), Mei4−/−, Spo11−/−, and Rec114−/− mice were immunoprecipitated with an anti-MEI4 antibody. Input extracts and immunoprecipitated fractions were probed with anti-REC114 antibody.
- Figure 4. MEI4, REC114 and IHO1 form a complex.
(A) Immunostaining of REC114 and SYCP3 in early prophase spermatocytes from Rec114+/+ (14 dpp), Mei4−/− (12 dpp), and Rec114−/− (14 dpp) males. Scale bar, 10 μm. (B) Quantification of REC114 foci in leptotene spermatocytes from 13 dpp Rec114+/+, Mei4−/−, Spo11−/−, and Rec114−/− males. The numbers (mean ± SD) of total foci and of foci on chromosome axis (co-localized with SYCP3) were: 245 ± 58 and 187 ± 47 for Rec114+/+ (n = 83 nuclei), 241 ± 68 and 189 ± 55 for Spo11−/− (n = 30), 13 ± 11 and 3 ± 3 for Rec114−/− (n = 69), and 32 ± 17 and 14 ± 11 for Mei4−/− (n = 52). P values were calculated with the Mann–Whitney two-tailed test. (C) Co-immunoprecipitation of REC114 and IHO1 with MEI4. Total testis extracts from 14 dpp Rec114+/+ (WT), Mei4−/−, Spo11−/−, and Rec114−/− mice were immunoprecipitated with an anti-MEI4 antibody. Input extracts were probed with anti-IHO1 and anti-SYCP3 antibodies. MEI4 is not detected in input extracts. Immunoprecipitated fractions were probed with anti-IHO1, anti-MEI4, and anti-REC114 antibodies.
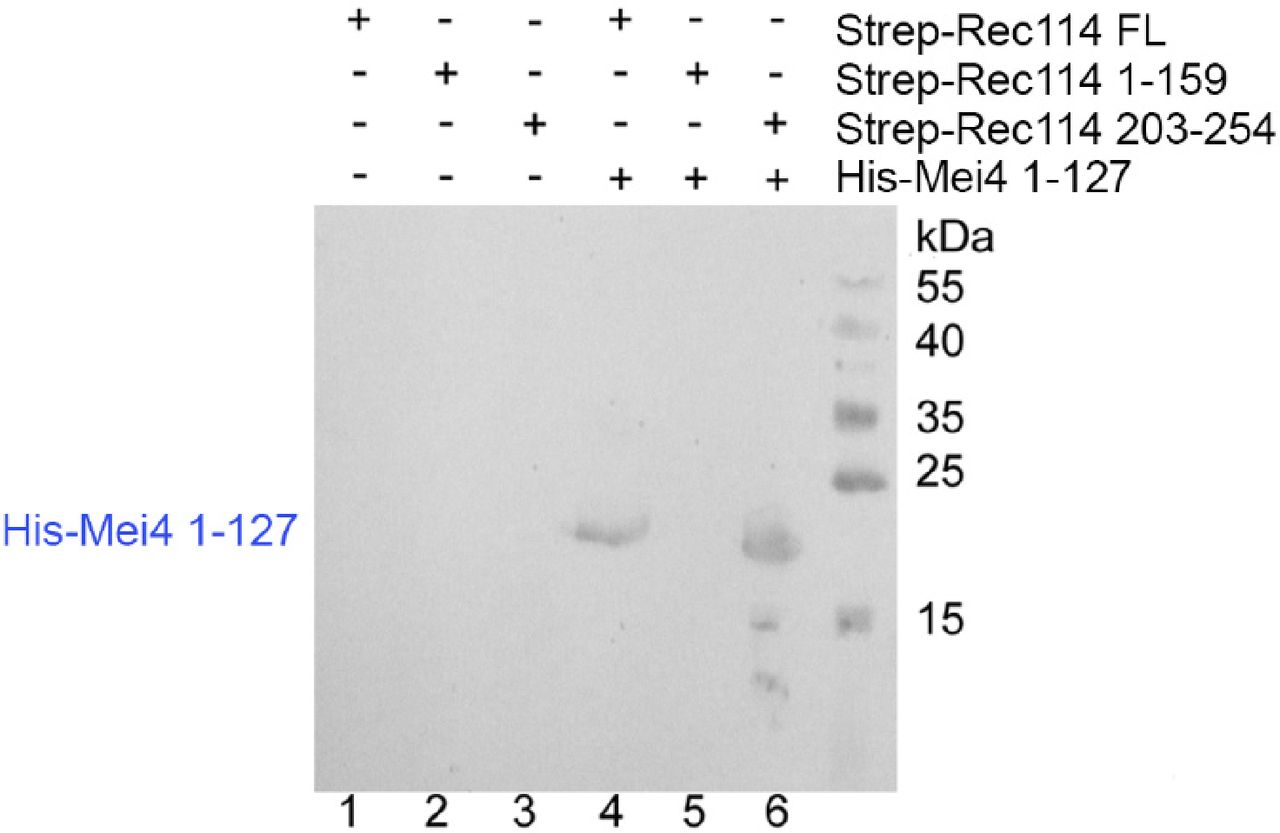
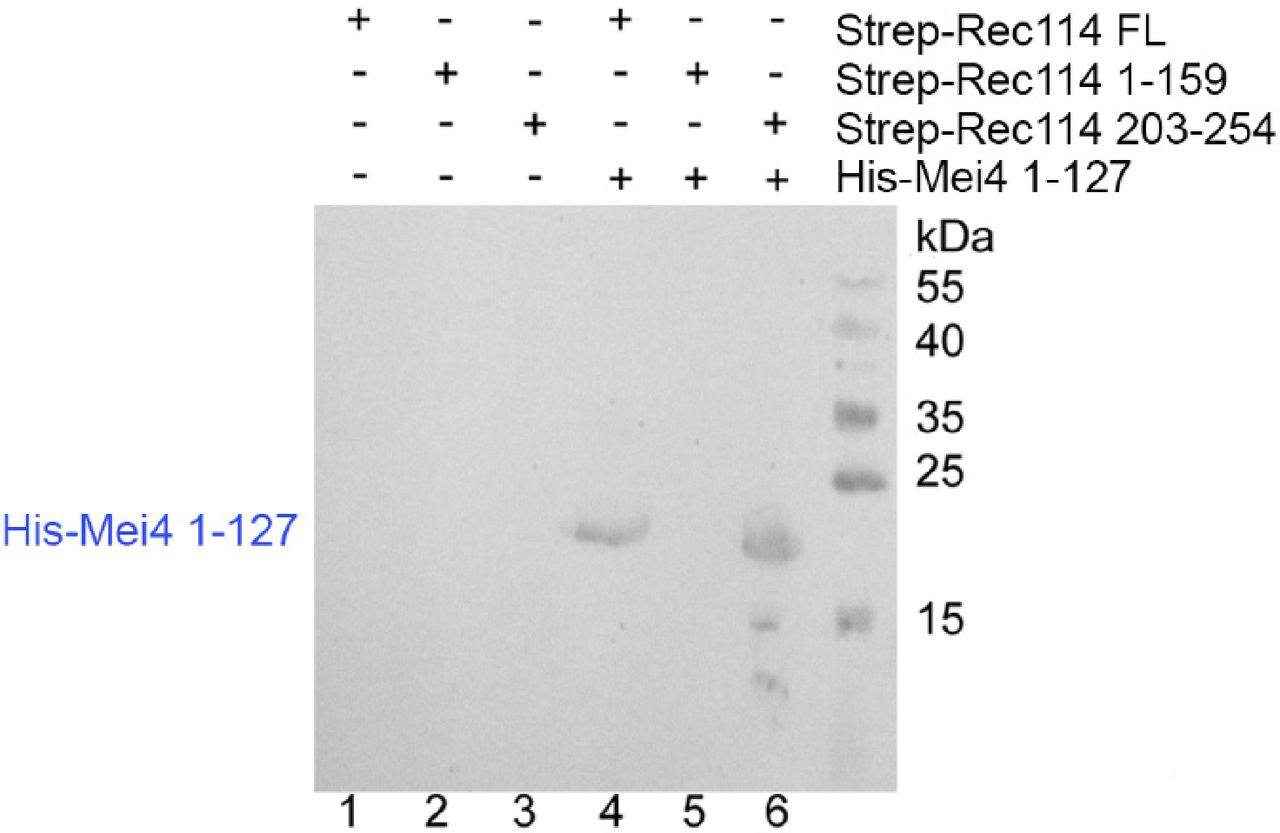
- Figure 5. REC114 forms a stable complex with MEI4.
A Strep-tag pull-down experiments with the REC114 and MEI4 constructs are shown in the upper panel. Full length and fragments of Strep-REC1114 were purified alone or after co-expression with MEI4 (1-127). Strep-REC114 (203-254) was insoluble on its own (lane 3), but when co-expressed with MEI4 (1-127) became soluble and was sufficient for interaction with MEI4 (lane 6). MEI4 (1-127, blue star) was pulled down by full-length (FL) REC114 and REC114 (203-254), but not by REC114 (1-159) (lanes 4–6). Proteins were detected by Coomassie blue staining. (A) Western blot confirmation of His-Mei4 with anti-His antibody is shown in Fig S10. Red star indicates a degradation fragment of FL REC114. (B) Strep-tagged REC114 (203-254) was co-expressed with His-MEI4 (1-127) and purified first using Strep-Tactin resin. The His-tag of MEI4 was removed by TEV protease cleavage followed by a passage through a Ni2+ column. The complex was then purified by Superdex 200 size-exclusion chromatography. The gel filtration elution profile is shown. (C) SDS–PAGE analysis of the peak fractions shown in (B). Proteins were detected by Coomassie blue staining.
- Figure S10. Western blot analysis of REC114/MEI4 affinity purification
Protein samples shown in Fig 5A were analyzed by Western blotting using an anti-His antibody confirming the presence of His-MEI4 1-127 (predicted molecular weight 17.4 kD) co-purifying with Strep-REC114 FL and 203-254 in lanes 4 and 6, respectively.
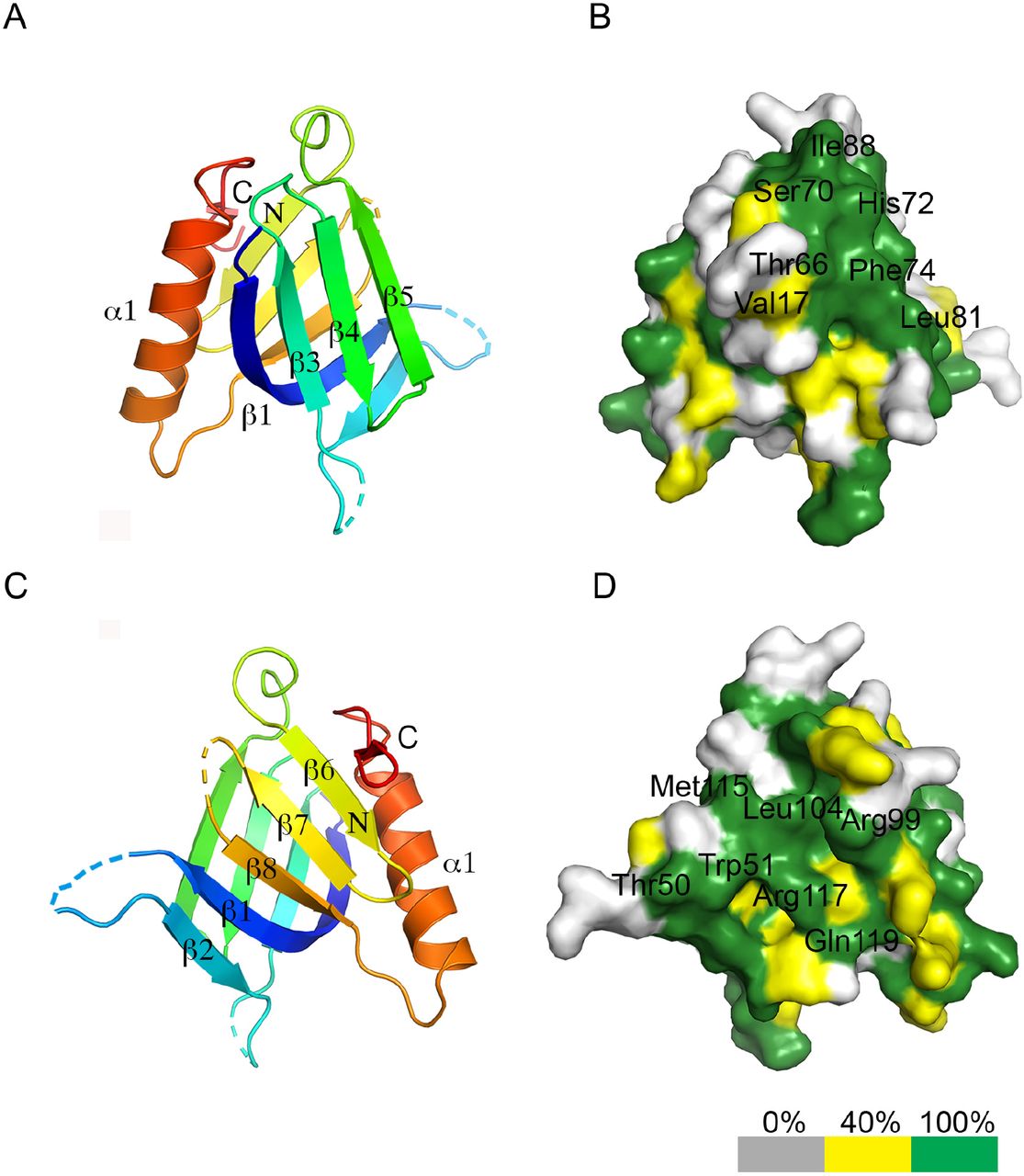
- Figure 6. Crystal structure of the REC114 PH domain.
(A) Schematic representation of the PH domain structure in mouse REC114, based on this work. (B) Ribbon diagram of the REC114 PH domain. The polypeptide chain is colored from the N-terminus (blue) to the C-terminus (red). Missing residues in the loops are shown by dashed lines. (C) The same ribbon diagram as in (A), but rotated 180° around the vertical axis. (D) Sequence alignment of REC114 proteins. Residues that are 100% conserved are in solid green boxes. The secondary structures of REC114 are shown above the sequences. The previously identified conserved motives, SSM1 to SSM6 (Kumar et al, 2010; Tesse et al, 2017), are shown below the sequence. Mouse, Mus musculus: NP_082874.1; Human, Homo sapiens: NP_001035826.1; Bos, Bos mutus: XP_005907161.1; Meleagris, Meleagris gallopavo: XP_019474806.1; Aquila, Aquila chrisaetos canadensis: XP_011595470.1; Xenopus, Xenopus laevis: OCT89407.1.
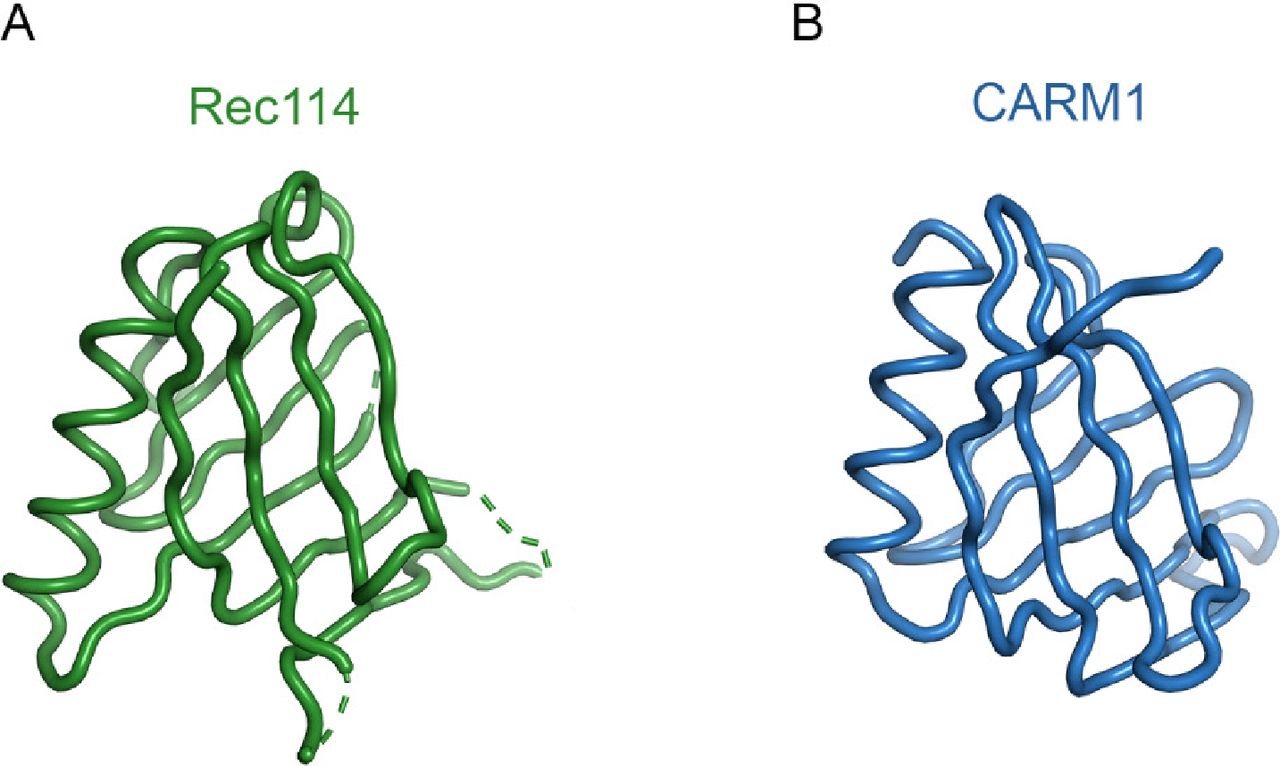
- Figure S11. Comparison of the REC114 PH domain with the N-terminal domain of CARM1.
(A) Structure of mouse REC114 (15-150). (B) Structure of CARM1 (2OQB) superimposed to REC114 with r.m.s. deviation of 1.69 Å for 87 Cα atoms.
- Figure S12. Conservation of the REC114 PH domain surface.
(A) Ribbon diagram of the REC114 PH domain. (B) Surface representation of the PH domain in the same orientation as in (A). The conservation of surface residues is represented from grey to green, according to the color scale shown below and based on the sequence alignment shown in Fig 6D. The positions of the most conserved and exposed residues are shown. (C) Ribbon diagram of the PH domain, but rotated 180° around the vertical axis, compared with (A). (D) Surface conservation of the PH domain corresponding to the view shown in (C).
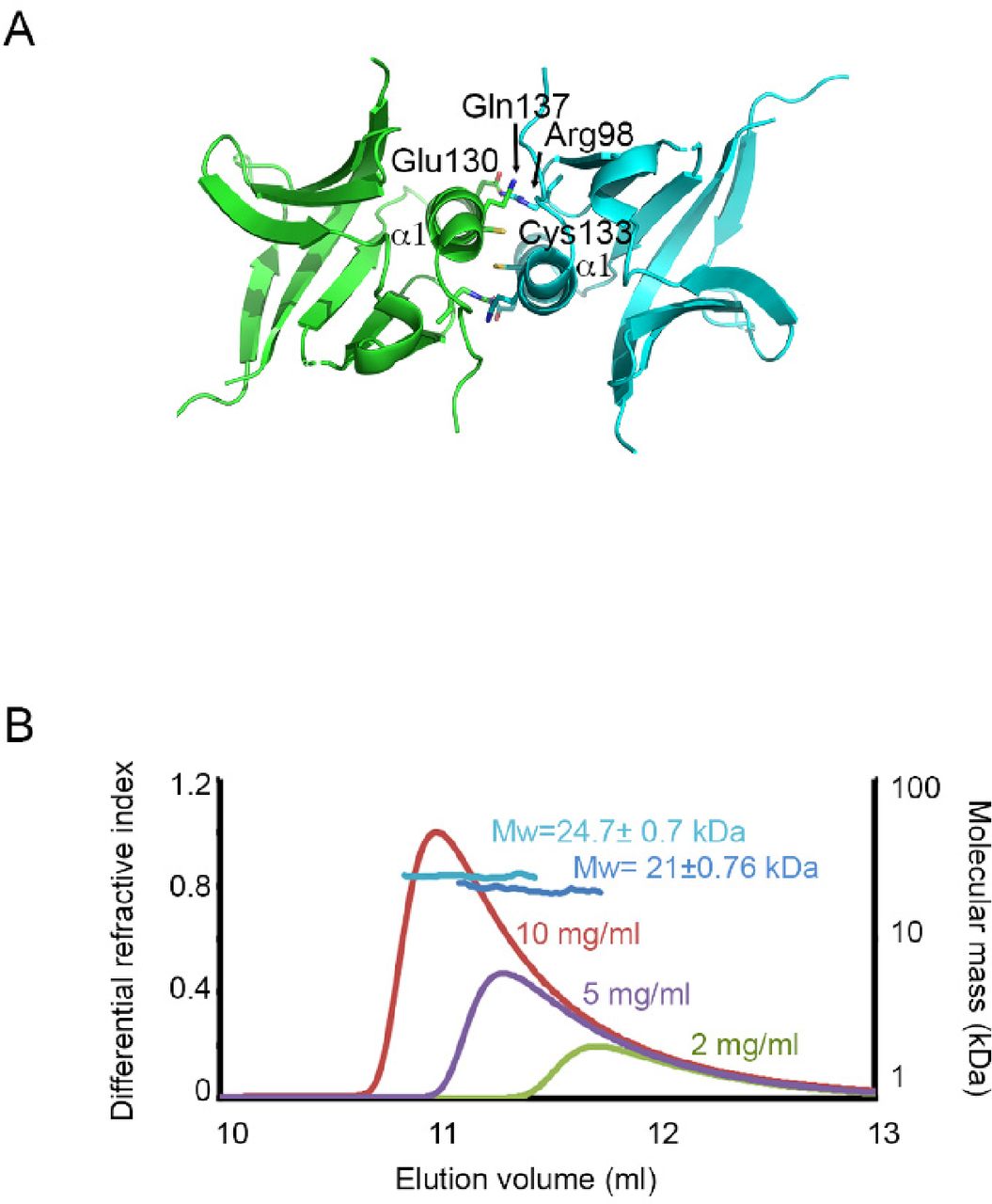
- Figure S13. Dimerization of Rec114.
(A) The REC114 PH domain forms dimers with a symmetry-related molecule. The dimer interface includes the α1 helix and β6 strand. The key interacting residues are indicated. (B) Molecular mass determination of the mouse REC114 PH domain by multi-angle laser light scattering (MALLS). 50 μl of purified protein at a concentration of 10, 5, and 2 mg/ml was injected in a Superdex 75 gel filtration column. Online MALLS and refractive index data were recorded on a DAWN-EOS detector (Wyatt Technology Corp.) using a laser emitting at 690 nm and an Optilab T-rEX detector (Wyatt Technology Corp.), respectively, with a refractive index increment dn/dc of 0.185 ml·g−1. Data were analyzed using the ASTRA 6 software (Wyatt Technology Corp.). The molecular mass could only be determined for samples injected at 10 and 5 mg/ml (MW in light and dark blue, respectively). The results indicate that REC114 exists in solution as a concentration-dependent mixture of dimers and monomers with a fast exchange rate.












Tables
REC114 native REC114 SeMet Data collection Space group P6122 P42212 Cell dimensions a, b, c (Å) 107.5, 107.5, 82.8 88.5, 88.5, 101.5 α, β, γ (°) 90, 90, 120 90, 90, 90 Resolution (Å) 93–2.5 (2.57–2.5)a 66.7–2.7 (2.8–2.7) Rmerge 5.7 (212.1) 12.2 (108.8) I/σI 24.2 (1.11) 11.1 (1.68) CC1/2 1 (0.548) 0.997 (0.725) Completeness (%) 99.6 (97.5) 100 (99.9) Redundancy 11.2 (11.1) 7.1 (7.1) Refinement Resolution (Å) 46.6–2.5 No. reflections 10,215 Rwork/Rfree 25 (30) B-factors 77.3 R.m.s. deviations Bond lengths (Å) 0.014 Bond angles (°) 1.641 ↵a Values in parentheses are for the highest resolution shell.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

In this Issue
Subjects
Related Articles
Cited By...
- Principles of chromosome organization for meiotic recombination
- Seeding the meiotic DNA break machinery and initiating recombination on chromosome axes
- Cryo-EM structure of the Spo11 core complex bound to DNA
- Essential roles of the ANKRD31-REC114 interaction in meiotic recombination and mouse spermatogenesis
- Evolutionary conservation of the structure and function of meiotic Rec114-Mei4 and Mer2 complexes
- Evolutionary conservation of the structure and function of meiotic Rec114-Mei4 and Mer2 complexes
- Structure and DNA-bridging activity of the essential Rec114-Mei4 trimer interface
- Essential roles of the ANKRD31-REC114 interaction in meiotic recombination and mouse spermatogenesis
- Structure and DNA bridging activity of the essential Rec114-Mei4 trimer interface
- Characterization of the REC114-MEI4-IHO1 complex regulating meiotic DNA double-strand break formation
- Evolutionary conservation of the structure and function of meiotic Rec114-Mei4 and Mer2 complexes
- TOPOVIBL-REC114 interaction regulates meiotic DNA double-strand breaks
- Evolution and diversity of the TopoVI and TopoVI-like subunits with extensive divergence of the TOPOVIBL subunit
- Phosphoregulation of DSB-1 mediates control of meiotic double-strand break activity
- Caenorhabditis elegans DSB-3 reveals conservation and divergence among protein complexes promoting meiotic double-strand breaks
- C. elegans DSB-3 Reveals Conservation and Divergence among Protein Complexes Promoting Meiotic Double-Strand Breaks
- The histone modification reader ZCWPW1 links histone methylation to PRDM9-induced double strand break repair
- Homozygous mutations in REC114 cause female infertility characterised by multiple pronuclei formation and early embryonic arrest
- DNA-dependent macromolecular condensation drives self-assembly of the meiotic DNA break machinery
- Ensuring meiotic DNA break formation in the mouse pseudoautosomal region