



Article Figures & Data
Figures
- Figure 1. DNMT3A variants cause unique DNAm signatures.
(A) Schematic representation of five protein-coding DNMT3A splice variants and their functional protein domains. The target sites of shRNA are indicated. (B, C) KD (B) and OE (C) of individual transcripts was confirmed with RT-qPCR (relative expression versus GAPDH; mean ± SD; n = 3). (D–F) Scatter plots of DNAm profiles upon down-regulation of transcripts 1+3 (D), transcript 2 (E), and transcript 4 (F) compared with scrambled shRNA controls (mean β-values; n = 3). Significantly hyper- and hypomethylated CpGs are depicted in red and blue, respectively (adj. P < 0.05). (G) Venn diagrams demonstrate overlap of DNAm changes upon modulation of transcripts 1+3 and transcript 2. (H–J) Scatter plots of DNAm profiles upon OE of transcripts 1+3 (H), transcript 2 (I), and transcript 4 (J), as compared with empty control vectors without any DNMT3A transcript (additional independent biological replicates, n = 3). Highlights indicate the CpGs that were significantly changed upon KD of the corresponding transcripts (hyper- and hypomethylated upon KD in red and blue, respectively; see Fig 1D and E). Almost all CpGs are modified in opposite directions upon KD and OE and the corresponding P-values are indicated (Fisher's t test). (K) Enrichment of histone marks within 250 bp up- and downstream of each relevant CpG with significant DNAm changes upon KD of transcripts 1+3, transcript 2, and random 50,000 CpGs (Background). Fold change (FC) was calculated over the input background signal. *P < 0.05, **P < 0.01, ***P < 0.001 (t test).
- Figure S1. Validation of DNMT3A splice variant expression.
(A) KD and (B) OE of DNMT3A transcripts (Tr.) by RT-qPCR relative to control samples scrambled and empty, respectively (KD and OE each in independent biological replicates, n = 3; mean ± SD). Significance was estimated by the t test; *P < 0.05, **P < 0.01, ***P < 0.001. (C) Western blot analysis of DNMT3A isoforms upon OE. Please note that the N-terminal antibody targets only transcripts 1+3 and 4 (expected at ∼130 kD and 18 kD, respectively). Actin was used as a reference protein. The Western blot was performed with limited cell numbers, and protein levels, therefore, differ between samples. Protein validation upon OE of Tr.2 and KD of all transcripts was not possible because of limited cell amount and unreliable antibody targeting the C-terminus (results not demonstrated).
- Figure S2. Cross-comparison of DNAm changes upon KD of DNMT3A splice variants.
(A) Hypermethylated CpGs upon KD of transcripts 1+3 (Tr.1+3; 25 CpGs) did not overlap with hypomethylated CpGs upon KD of transcript 2 (Tr.2; 369). (B) Furthermore, the overlap was extremely low when the analysis was performed vice versa.
- Figure S3. Enrichment of chromatin marks around differentially methylated CpGs.
We utilized ChIP-seq of short-term cultured human CD34+ cells from the IHEC. Enrichment of histone marks within 250 bp up- and downstream of CpGs with significant DNAm changes upon KD of transcripts 1+3 (A) and transcript 2 (B). The black lines exemplarily depict enrichment around random 50,000 CpGs (background). Enrichment often peaked around the differentially methylated CpGs.
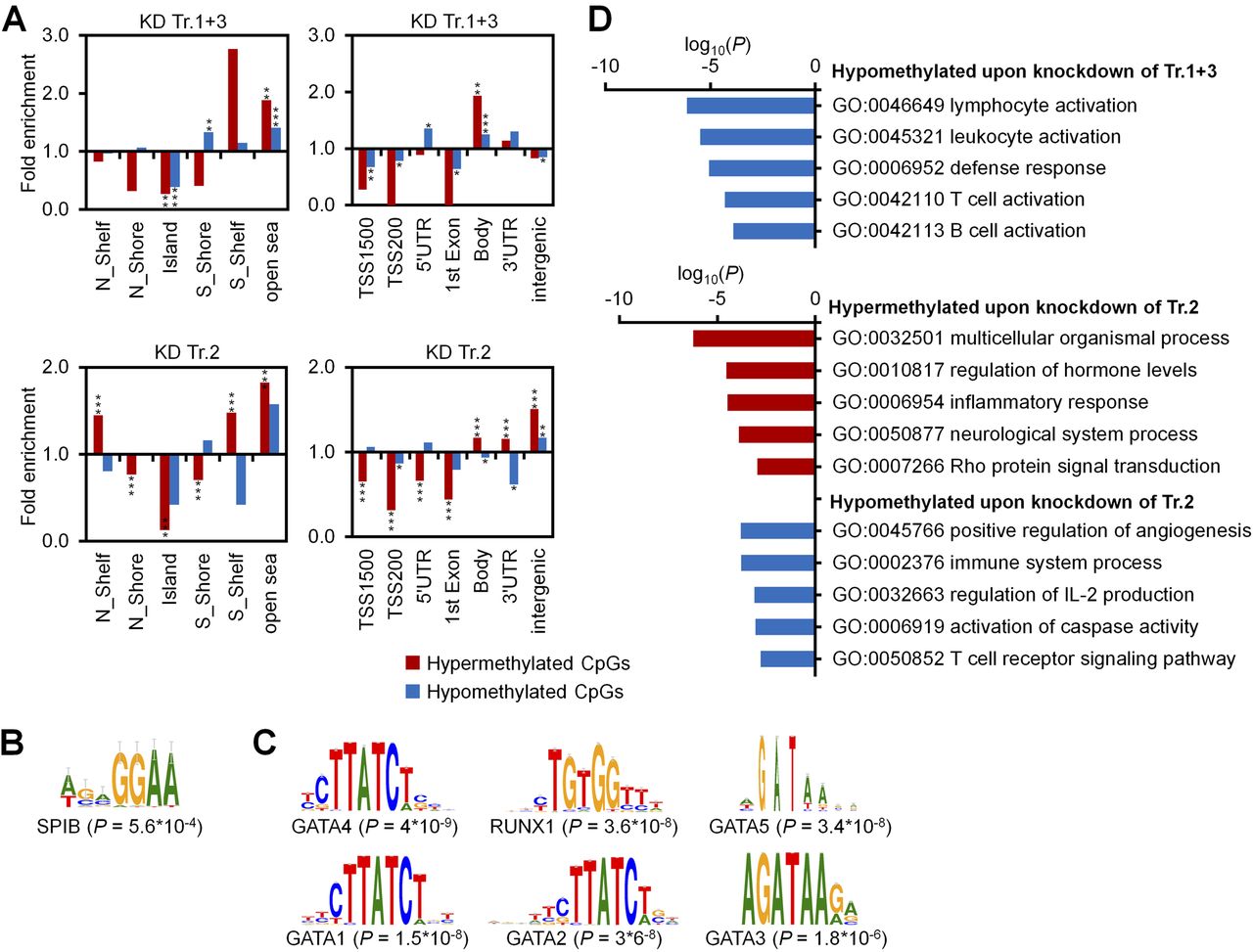
- Figure S4. Enrichment analysis of differentially methylated CpGs.
(A) Enrichment analysis of significant differentially methylated CpGs upon KD of transcripts 1+3 (25 hyper- and 327 hypomethylated CpGs) and of transcript 2 (8,536 hyper- and 369 hypomethylated CpGs) as compared with all CpGs on the microarray in relation to CGIs (left panels) and gene regions (right panels). Two kb upstream or downstream of CGIs were termed as shore region (“north” [N] and “south” [S], respectively); 2 kb flanking regions next to the shores were defined as shelf regions; and all other CpG sites were referred to as “open sea”; TSS1500 and TSS200 = 1,500 and 200 bp upstream of transcription start site, respectively. Significance was calculated by hypergeometric distribution; *P < 0.05, **P < 0.01, ***P < 0.001. (B, C) Motive enrichment analysis of DMRs (flanking 50 bp of significant CpGs) for KD of transcripts 1+3 (B) and transcript 2 (C). (D) GO classification of genes with significant DNAm changes upon KD of transcripts 1+3 and transcript 2.
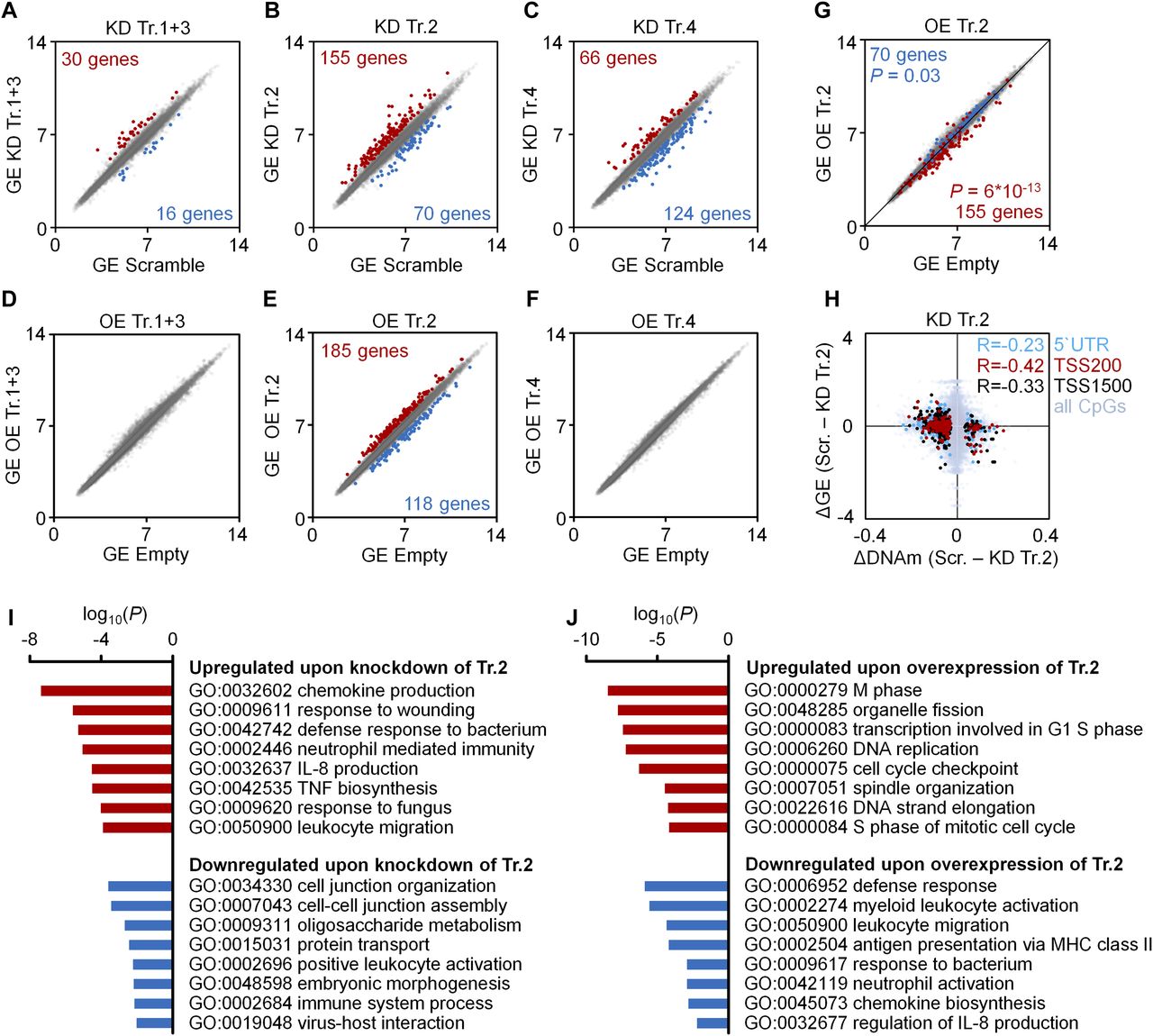
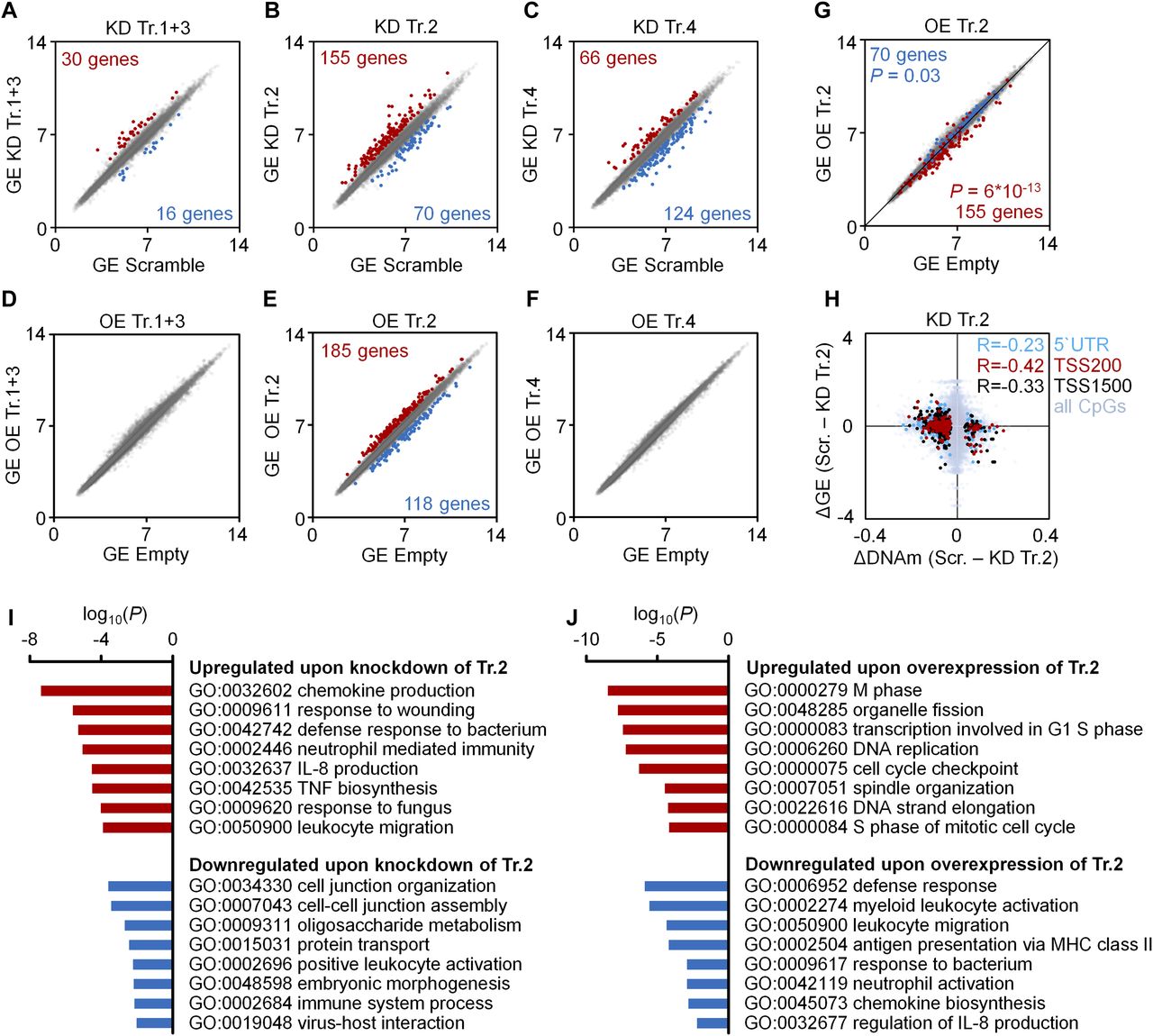
- Figure 2. Gene expression changes upon modulation of DNMT3A transcripts.
(A–F) Scatter plots of gene expression (GE; Affymetrix Gene ST 1.0 microarray) upon KD (A–C) or OE (D–F) of transcripts 1+3 (A, D), transcript 2 (B, E), and transcript 4 (C, F), as compared with scrambled shRNA controls or empty control vectors (mean log2 values of normalized data; n = 3). Numbers of genes that reached statistical significance (adj. P < 0.05) and at least 1.5-fold up- or down-regulation are indicated in red and blue, respectively. (G) To visualize that gene expression changes upon KD and OE of transcript 2 occurred preferentially in opposite directions, the scatter plot depicts significant down-regulation (blue) and up-regulation (red) upon KD of transcript 2 in the data for OE of transcript 2 (Fischer's t test). (H) DNAm changes of CpGs upon KD of transcript 2 were matched to differential expression of the corresponding genes. Hypermethylated CpGs in promoter regions were associated with down-regulation of gene expression and vice versa (R = −0.23, R = −0.42 and R = −0.33, respectively; n = 3). (I, J) GO analysis was performed to classify gene expression changes upon KD (I) and OE (J) of transcript 2.
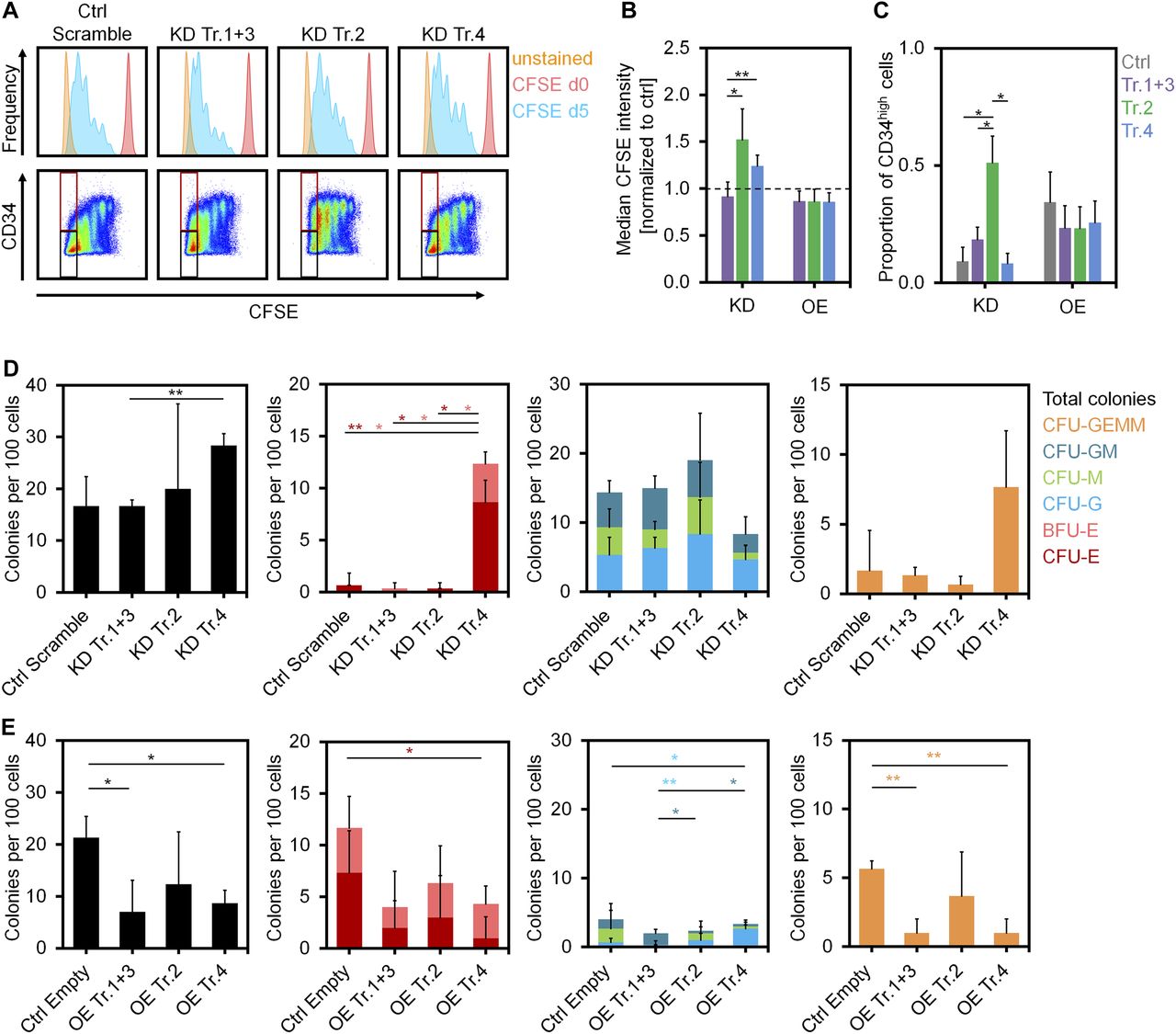
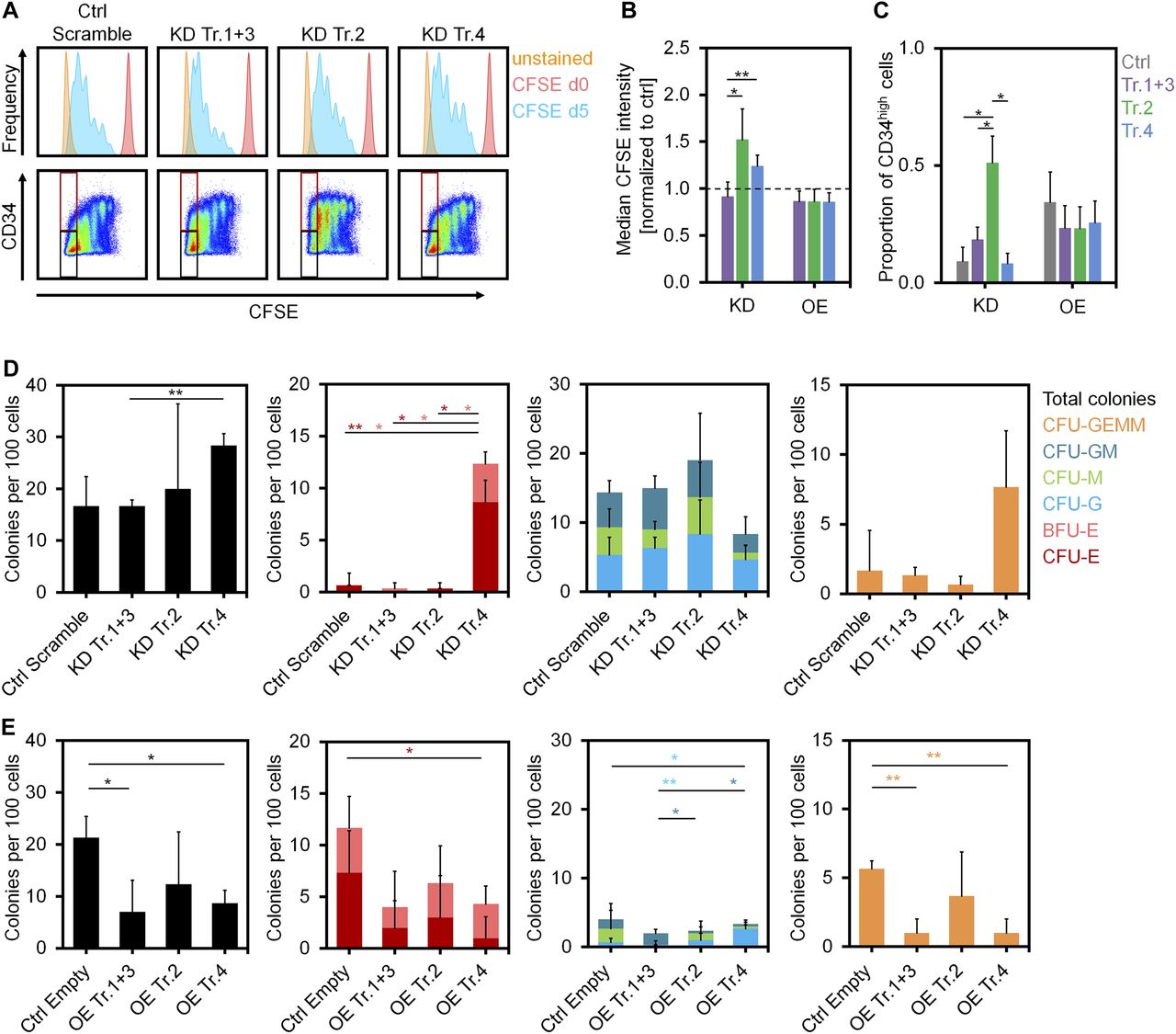
- Figure 3. Impact of DNMT3A variants on proliferation and differentiation of HSPCs.
(A) Histograms of residual CFSE staining to estimate proliferation of CD34+ cells after transfection with shRNAs after 5 d (blue). For comparison, measurements at day of HSPC isolation (day 0, no cell division, shown in red) and unstained controls at day 5 (shown in orange) are provided. (B) KD of transcripts 2 and 4 resulted in higher CFSE retention (slower proliferation) than the control (dashed line; mean ± SD; n = 3). (C) The proportion of CD34 high cells in the fast proliferating fraction was increased upon KD of transcript 2 (gates are indicated in Fig 3A; mean ± SD; n = 3). (D, E) CFU frequency (per 100 initially seeded HSPCs) was analyzed after KD (D) and OE (E) of DNMT3A transcripts (mean ± SD; n = 3). *P < 0.05, **P < 0.01 (t test).
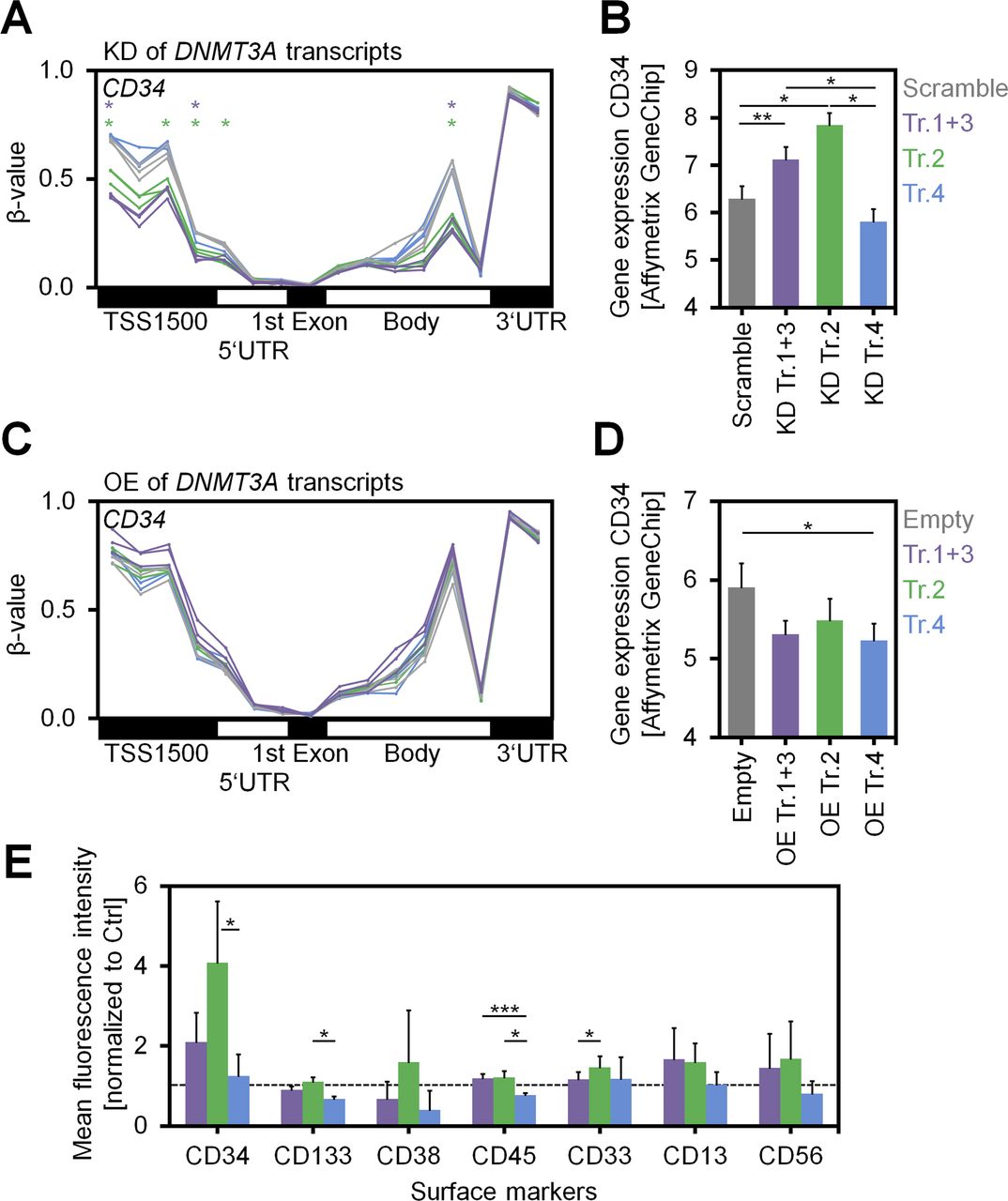
- Figure S5. Analysis of CD34 upon modulation of DNMT3A variants.
(A) DNAm levels (β-values) of CpGs associated with the gene CD34 reflect demethylation of the promoter region (particularly TSS1500) upon KD of transcripts 1+3 and transcript 2. Individual CpGs that reached statistical significance (adj. P < 0.05) are indicated by asterisks. (B) Gene expression of CD34 (Affymetrix Gene Chip; log2 signal intensities) was up-regulated upon KD of transcripts 1+3 and particularly upon KD of transcript 2 (n = 3; mean ± SD). (C, D) A complementary tendency of (C) promoter methylation and (D) gene expression of CD34 was observed upon OE of DNMT3A variants. (E) Immunophenotypic analysis of HSPCs with KD of the variant transcripts upon culture for 7 d (additional independent biological replicates, n = 3; mean ± SD). The mean fluorescence intensities were normalized to controls with scrambled shRNAs. In these independent replicates, CD34 was again up-regulated upon KD of transcripts 1+3 and transcript 2, whereas for other surface markers only moderate changes were observed. *P < 0.05, **P < 0.01, ***P < 0.001.
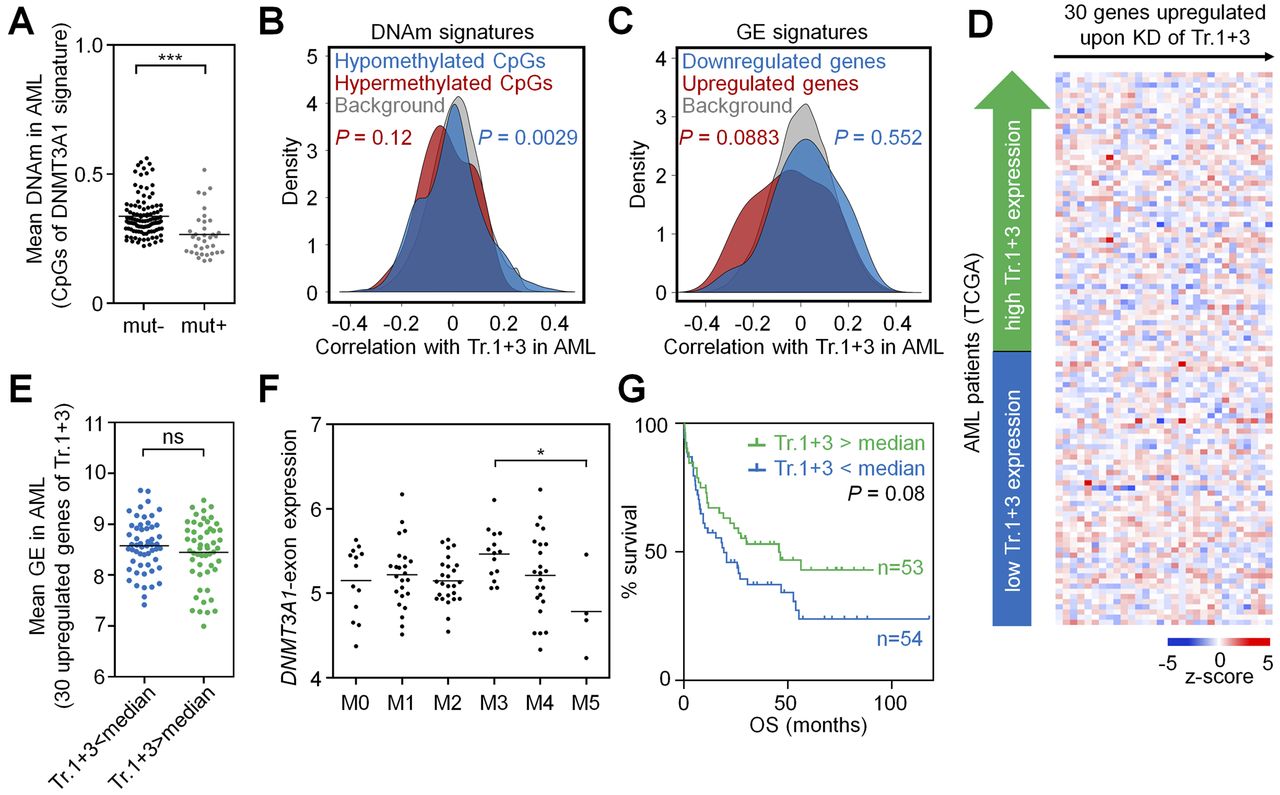
- Figure 4. DNMT3A2 signatures are coherently modified in AML.
DNMT3A2-associated DNAm and gene expression signatures are recapitulated in the AML dataset of TCGA (The Cancer Genome Atlas Research Network, 2013). (A) CpGs that revealed significant DNAm changes upon KD of transcript 2 in HSPCs in vitro (8,905 CpGs, see Fig 1E) had overall significantly lower mean DNAm levels in AML samples with a DNMT3A mutation—therefore, samples with DNMT3A mutation were excluded from further analysis. (B) As a surrogate for transcript 2 expression we used the transcript-specific DNMT3A2-exon (ENSE00001486123). CpGs that were either hypo- or hypermethylated upon KD of transcript 2 in HSPCs in vitro, revealed overall highly significant correlation with expression of the DNMT3A2-exon in AML. (C) In analogy, genes that were differentially expressed upon KD of transcript 2 in HSPCs in vitro were significantly related to expression of the DNMT3A2-exon in AML patients. (D) Heat map for association of DNMT3A2-exon expression in AML with expression of 155 genes that were up-regulated upon KD of transcript 2 in HSPCs. (E) These 155 genes were on average significantly higher expressed in AML patients with lower DNMT3A2-exon expression (stratified by median). (F) The DNMT3A2-exon is significantly lower expressed in the AML subgroups M4 and M5 (FAB-classification). (G) Kaplan–Meier plot indicates that lower DNMT3A2-exon expression (stratified by median) is associated with shorter OS. *P < 0.05, **P < 0.01, ***P < 0.001 (Mann–Whitney test).
- Figure S6. DNMT3A1 signatures are not recapitulated in AML.
DNMT3A1-associated DNAm and gene expression signatures are not recapitulated in the AML dataset of TCGA (The Cancer Genome Atlas Research Network, 2013). (A) CpGs that revealed significant DNAm changes upon KD of transcripts 1+3 in HSPCs in vitro (352 CpGs, see Fig 1D) had overall significantly lower mean DNAm levels in AML samples with a DNMT3A mutation—therefore samples with DNMT3A mutation were excluded from further analysis. (B) As a surrogate for transcript 1+3 expression, we used the transcript-specific DNMT3A1-exon (ENSE00001486208). CpGs that were either hypo- or hypermethylated upon KD of transcripts 1+3 in HSPCs in vitro revealed only moderate correlation for the hypomethylated CpGs with expression of the DNMT3A1-exon in AML. (C) In analogy, genes that were differentially expressed upon KD of transcripts 1+3 in HSPCs in vitro did not correlate to expression of the DNMT3A1-exon in AML patients. (D) Heat map for association of DNMT3A1-exon expression in AML with expression of 30 genes that were up-regulated upon KD of transcripts 1+3 in HSPCs. (E) There were no differences in the expression of these 30 genes between AML patients expressing lower or higher DNMT3A1-exon (stratified by median). (F) DNMT3A1-exon is not clearly correlated with the French–American–British classification. (G) Kaplan–Meier plot indicates that AML patients expressing low DNMT3A1-exon (stratified by median) have a tendency for shorter OS. *P < 0.05, **P < 0.01, ***P < 0.001 (Mann–Whitney test).
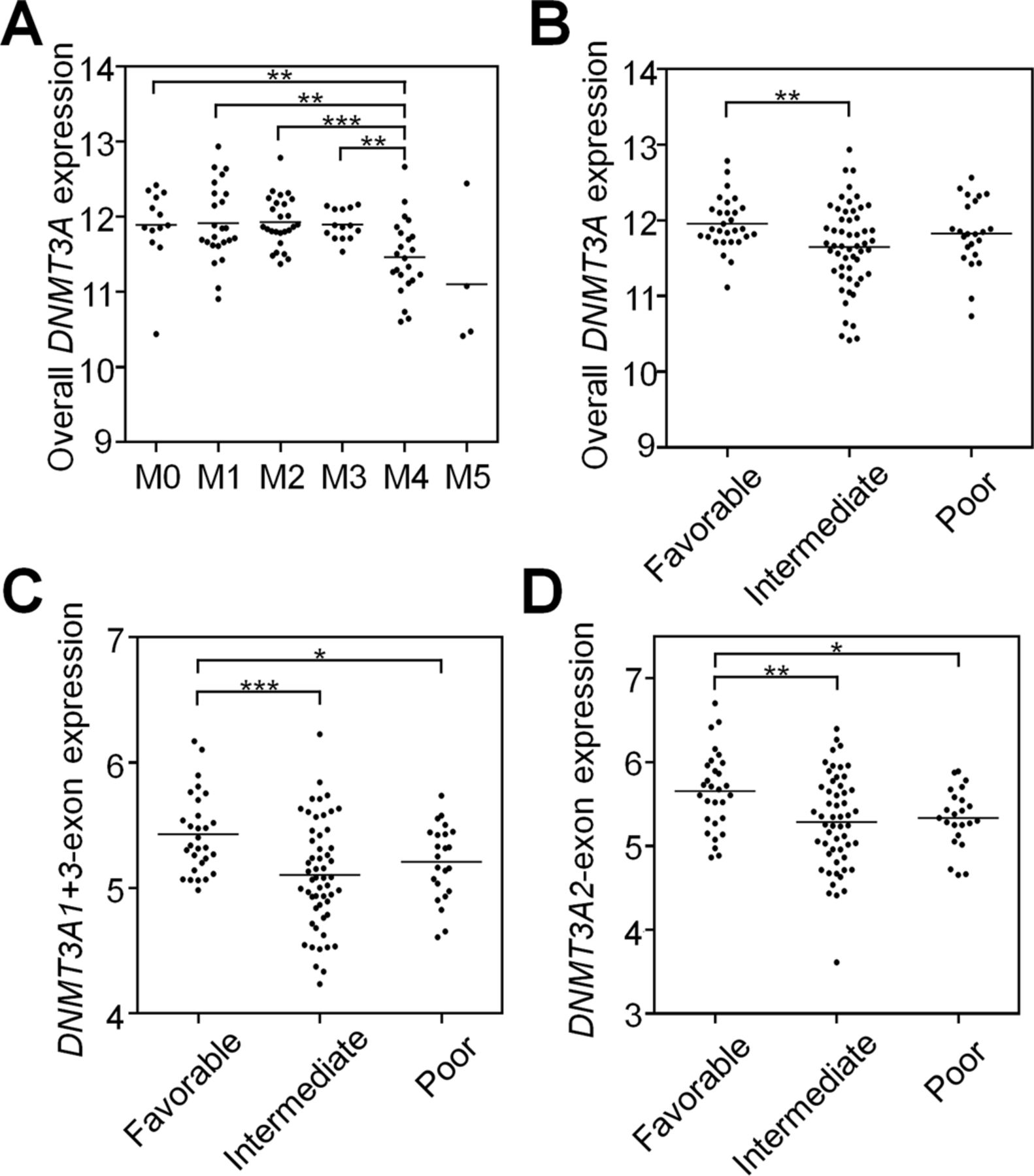
- Figure S7. Correlation of DNMT3A expression and its variants with clinical parameters.
Gene expression analysis was performed using the RNA–seq data of 107 AML patients from the TCGA repository. (A) Overall, DNMT3A expression correlated with the M4 group of the French–American–British classification and (B) with the intermediate cytogenetic risk group. Correlation of the DNMT3A transcript-specific exons (C) DNMT3A1-exon (ENSE00001486208) and (D) DNMT3A2-exon (ENSE00001486123) with cytogenetic risk groups. Statistical significance was estimated with the Mann–Whitney test. *P < 0.05, **P < 0.01, ***P < 0.001.







Supplementary Materials
Table S6 Transcript-specific RT-qPCR primers.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
